|
Az UNESCO minden évben egy kiemelkedő tudóst nevez
meg az „év emberének”. 2012-ben tudományos teljesítménye
elismeréseként a száz éve született Szentágothai János lett az „év
embere”. A kiváló anatómus tudományos munkássága, az idegrendszer
funkcionális szerkezetének kutatása során igen kiterjedt témakörben
ért el világviszonylatban is kiemelkedő eredményeket. Szentágothai az
idegrendszeri szerkezetekben mindig a szerkezetben rejlő működést, s
megfordítva, az idegi működések alapjául szolgáló struktúrát kutatta.
Érdeklődése széles körű volt: az agykéreg, a látórendszer, az
agytörzs, a gerincvelő és a kisagy funkcionális szerkezetének kutatása
során ért el máig érvényes eredményeket. Szentágothai és munkatársai a
kisagy neurális szerkezetének elemzése során elért eredményei jelentős
mértékben járultak hozzá ezen agyrész működésének tisztázásához.
Kisagy
A cerebellum vagy „kisagy” neve ellenére igen fontos régiója az
agynak. Érdemes megemlíteni, hogy ez a viszonylag kis méretű, kis
volumenű (1. ábra) agyi régió az összes idegsejtek mintegy felét, kb.
százmilliárd idegsejtet tartalmaz. Ez is arra utal, hogy „kicsinysége”
ellenére sok fontos funkciót lát el. Ezek közül már régen ismert a
motoros funkciók koordinációjában játszott szerepe. Ma már tudjuk,
hogy a kisagy a motoros koordináción kívül számos más tevékenységben
is részt vesz, például a motoros tanulási folyamatban, sőt a kognitív,
a beszéddel, tudati jelenségekkel kapcsolatos működésekben is. Az
emberi kisagy, jóllehet nem életfontosságú, mégis sine qua non minden
emberi tevékenység számára a már említett motoros működésektől kezdve
a beszéden, egyes tudati tulajdonságokon keresztül egészen az
intellektuális képességekig (Eccles et al., 1967).
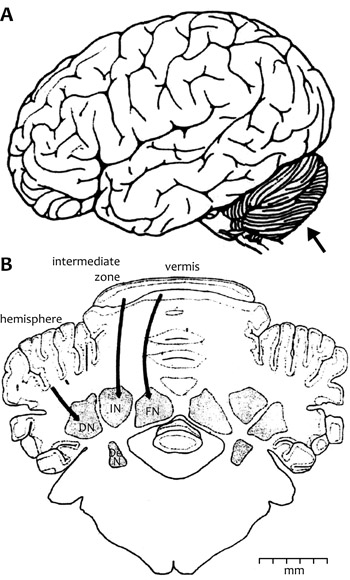
1. ábra • (A) (nyíl) – az agy hátsó részében
található • (B) a kisagy keresztmetszete: a kéreg-foliák
és a belső magvak között vastag nyilak jelzik
az összeköttetést. A jelzett kisagyi magok közül
legjelentősebb az oldalsó (DN).
A cerebellum törzsfejlődésileg az agy egyik legősibb, morfológiailag
pedig talán a legjellegzetesebb állománya (Hámori, 2012). A
kisagykéreg szerkezete ugyanis a halaktól az emberig, azaz az elmúlt
450 millió év során alig változott. A kisagy makroszkópos anatómiája a
törzsfejlődés során ugyan jelentősen módosult, de ez is inkább
mennyiségi, mint minőségi változás volt: a fejlődés során történt
térfogat-növekedés ugyanis a kéreg szaporulatát a kisagyi magvak [a
kisagynak a kéreg mellett a másik, fontos állománya (1. ábra)] hasonló
fejlődése kísérte. Így az arány e két fő komponens, a kéreg és a kéreg
alatti kisagyi magok között lényegében nem változott, bár maga a
kisagy, főleg az emlősöknél, igen nagyra nőtt.
A kisagy mikroszkópos anatómiája
Külön tárgyaljuk a rétegezett kérget és a belső fehérállományba
ágyazott kisagyi magvakat.
A.) Kisagykéreg
A kisagykéreg külső, háromrétegű szürkeállományból áll, amely a
vékony, rostokat tartalmazó fehérállományt borítja. A kéreg jól
látható redőzöttségét az egymásra hajtogatott, haránt irányú foliumok
okozzák. A foliumok (1. ábra) a fehérállomány központi részéből
kiindulva szinte „szétsugároznak”, s így, longitudinális (szagittális)
metszetben a kisagynak a Thuja occidentalis-ra emlékeztető formáját
alakítják ki, amelyet korábbi anatómusok igen szemléletesen az „élet
fájának” (arbor vitae) neveztek el.
A három réteg:
1.) szemcsés réteg, melyet a fehérállomány határol,
2.) a legfelületesebb, pia mater által határolt molekuláris réteg, és
3.) a két réteg között elhelyezkedő, egysejtrétegű Purkinje-sejt (vagy
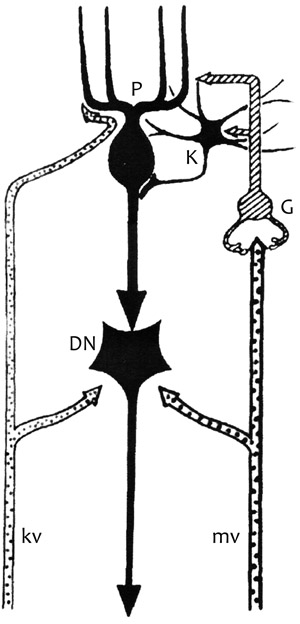
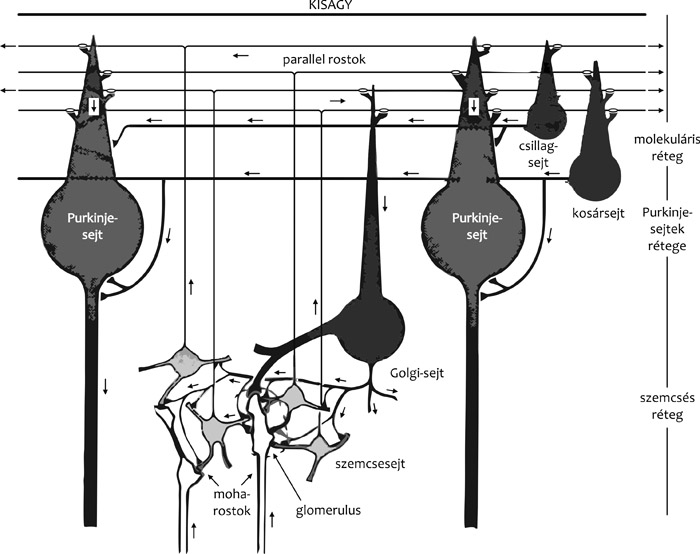
ganglionáris réteg) (2. ábra).
1.) Szemcsés réteg • Nevét a rendkívül szorosan egymáshoz szorított
kicsiny idegsejtektől, a szemcsesejtektől nyerte. Ezek az agy
legkisebb idegsejtjei (5–8 mm), számuk azonban óriási. Az átlagban 133
grammos emberi kisagy 100 milliárd idegsejtjéből 94 milliárd
szemcsesejt. A szemcsesejtnek három-öt rövid dendritje van (2. ábra),
melyek végei fogazottak, és több kis, rákollószerű, ún. protrúzióval
csatlakoznak a preszinaptikus moha axonokhoz. A szemcsesejt axonja
általában csupasz, nem mielinhüvelyes, és a molekuláris rétegbe jutva
„T”-alakban elágazódik, majd a sok („parallel”) axon kötegekbe
rendeződve fut a Purkinje-sejtek dendritfájára merőlegesen, a foliumok
hossztengelyében. Az axonok hossza a folium hosszméretétől függően 2–7
mm lehet (2. ábra). (Eccles et al., 1967).
A kis méretű, „serkentő” szemcsesejtek mellett elszórtan nagyobb
sejttestű, ún. Golgi-, gátló idegsejtek találhatók.
A kisagyi glomerulus vagy kisagyszigetek (2. ábra)
sejtmentes
területek a szemcsesejtek közé ékelve (Hámori – Szentágothai, 1966). A
glomerulus fő elemei:
a.) szemcsesejt dendritek az ollószerű végződésekkel,
b.) serkentő (glutamaterg) moharost végződés és
c.) gátló, GABA-erg axonok. Mindezen pre- és posztszinaptikus elemek
igen bonyolult, komplex szinaptikus szerveződést hoznak létre.
A rendszerint centrális helyszetű moharost végződés általában
serkentő. A moharost a dendritekkel aszimmetrikus szinaptikus
érintkezéseket létesít. A kisebb, varikózus, a glomerulus perifériáján
található Golgi-axonvégződés minden esetben gátló. A szemcsesejt
dendritek térbeli rekonstrukciójával kiderült, hogy egy glomerulusban
kb. 54 szemcsesejt dendrit található – mindegyik másik szemcsesejtből!
Minthogy minden dendritnek 3–5 posztszinaptikus végbunkója van, ezeken
keresztül egy-egy dendrit ugyanennyi szinaptikus kontaktust létesíthet
a moharosttal, a Golgi-axonnal vagy akár mindkettővel. Ennek
megfelelően a moharost átlagban 145, míg a Golgi-axon 87 szinaptikus
kontaktust létesít a szemcsesejt dendritekkel, azaz összességében
(átlagban) 232 szinaptikus érintkezés található egy kisagyi
glomerulusban. Ezek a számok is érzékeltetik, hogy a glomerulus
cerebelliben rendkívül komplex ingerületátadási folyamatok mennek
végbe, amelyekben a serkentő moharost szinapszisok és a gátló
Golgi-axon által kialakított szinapszisok működésének szimmetriája
vagy aszimmetriája határozza meg a posztszinaptikus szemcsesejtek (s
ezen keresztül a parallel axonok) ingerületi állapotát. Ez tovább
bonyolódik azzal, hogy a szemcsesejt négy dendritje négy különböző
moharosttól kap ingerületet, azaz a folyamatban az ingerdivergencia (1
moharost → 54 dendrithez a glomerulusban), illetve konvergencia (4
különböző moharost → 1 szemcsesejt) egyaránt jelentős szerepet
játszik.
2.) Purkinje-sejtek rétege • A Purkinje-sejteket, a központi
idegrendszer talán legkarakterisztikusabb idegsejtféleségét Jan
Evangelista Purkyne (1787–1869), a Breslauban, majd Prágában működő
cseh élettan- és szövettanprofesszor látta meg először, ezért róla
nevezték el. A nagy, ovoid Purkinje-sejttestek (16–30 × 21–40 mm) a
szemcsés réteg és a felületes, molekuláris réteg közé ékelt
egysejtréteget alkotnak. A Purkinje-sejtek fő dendritnyúlványa a
molekuláris rétegbe száll fel, s a szagittális síkban többszörösen
osztódva a kéreg felszínéig húzódik, míg axonja a sejttest másik
pólusán eredve a szemcsés rétegen keresztül (ahol mielinizálódik) éri
el a fehérállományt. Az axon induló része, vagyis a nem velőhüvelyes
szakasz 15–20 mm hosszú. A viszonylag vastag mielinhüvelyes rostok
többnyire a kisagyi magvakhoz futnak, s e magok fő gátló bemenetét
képezik (Eccles et al., 1967). Ito Maszao (Masao Ito) japán tudós
mutatta ki ugyanis, hogy a Purkinje-sejtek, illetve axonjaik gátlók,
és transzmitterük a GABA.
3.) Molekuláris réteg • A kisagykéreg elegáns, szemet gyönyörködtető
mértani elrendeződésének két fő tényezője a molekuláris rétegben
található. Ezek egyike a Purkinje-sejtek legyezőszerűen, szigorúan a
paraszagittális síkban szétterülő dendritfája, a másik az erre
pontosan merőlegesen (tehát a folium hossztengelyében) futó több
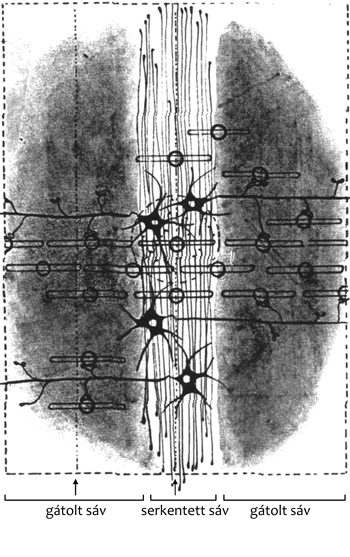
százezer parallel axon (2., 3., 4. ábra). A réteg három másik lényeges
neuronális eleme a két (gátló, GABA-erg) interneuron, a kosár- és
csillagsejt, valamint az olivo-cerebelláris pálya itt végződő axonjai,
a kúszórostok (2., 5. ábra).
A Purkinje-sejtek dendritfája a folium hossztengelyére merőleges,
szagittális síkban ágazódik el. E kétdimenziós arborizáció vastagsága
9–18 mm között van. A vastagabb elsődleges és másodlagos dendriteken
viszonylag kevés dendrikus tüske található, míg a harmadlagos vékony
dendritek, az ún. tüskés dendritek viselik egy Purkinje-sejt 30 000-80
000 posztszinaptikus tüskéinek túlnyomó többségét (3. ábra). (Eccles
et al., 1967)
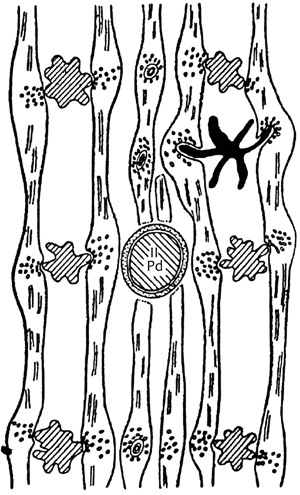
3. ábra • Az átkeresztezéses (crossing over) szinaptikus rendszer a
kisagykéreg molekuláris rétegében. • Pd – Purkinje fődendrit. A kisebb
dendritek tüskéi szinaptizálnak a dendritek között futó parallel
axonokkal. Fekete tüskés dendrit: kosársejt dendritje.
A vastagabb (elsődleges, másodlagos) dendriteken általában három
különböző axon végződik. Két végződéstípus GABA-t tartalmaz, ami a
gátló szinapszisokra jellemző. E két gátló végződést a kosár- és
csillagsejtek axonjaiként azonosították. A harmadik fajta
axonvégződés, a kúszórost szinapszisa excitatórikus. A kúszórostok
eredetét az egyik agytörzsi magban (oliva inferior) Szentágothai János
és Rajkovics Károly (1959) kísérletes módszerek segítségével még az
50-es években azonosította, s így értelmezhetővé tette a kisagy két fő
afferensének (moharostok, kúszórostok) a kisagy működését alapvetően
befolyásoló funkcionális szerepét. A kúszórostok mielinhüvelyüket a
Purkinje-sejtek magasságában veszítik el. A velőtlen rost szorosan
hozzáfekszik a Purkinje-sejt dendritfájának vastagabb ágaihoz, annak
oszlásait is követve, s a hosszú érintkezés során számos, egyes
számítások szerint 400–500 aszimmetrikus, serkentő szinaptikus
kontaktust létesít egy Purkinje-sejt, főként elsődleges és másodlagos
dendritjeivel. A szinaptikus kontaktusokat a kúszórost általában a
dendritek elágazódásánál vagy ahhoz közel hozza létre, méghozzá a
vastagabb dendritek öt-hat dendritikus tüskéjével. Ezek a tüskék tehát
– melyek a harmadlagos dendritek tüskéinél jóval rövidebbek – a fő
helyei a kúszórost – Purkinje-sejt közötti, rendkívül hatékony
excitatorikus szinaptikus transzmissziónak. Figyelemre méltó, hogy bár
az aszparaginsav volt a kúszórost lehetséges transzmitterei között az
első számú jelölt, a posztszinaptikus receptorok közül a metabotrop
glutamát receptor dendritikus tüskékben való erőteljes jelenléte
valószínűvé teszi azt, hogy egyedül vagy az aszparaginsavval együtt a
glutaminsav működik transzmitterként a kúszórost–Purkinje-sejt
szinapszisban.
Jellemző a már Santiago Ramón y Cajal által is
felismert kvantitatív viszony kúszórost és Purkinje-sejt között. Egy
Purkinje-sejtet egy kúszórost idegez be, mint ahogy említettük,
400–500 egyedi
|