|
|
Ö sszefoglalás
Darwin evolúciós elméletének ökológiai magva a létért
folyó küzdelem elkerülhetetlenségének gondolata. Egy sor kiemelkedő
tudós Ernst Haeckeltől a versengést szisztematikus kísérletekkel
vizsgáló grúz Georgij Francsevics Gausén és a közlegelő tragédiájának
megfogalmazásáról híres Garrett Hardinon át a niche-elméletet
először formalizáló George Evelyn Hutchinsonig, az ökológiát „a létért
folyó küzdelem” tudományaként fogta fel. Ez a felfogás a nyolcvanas
évektől kezdve háttérbe szorult. Az idei kerek Darwin-évfordulók
kapcsán megmutatjuk, hogy a szelekció eredeti elmélete
újrafogalmazható a strukturált populációk, az adaptív dinamika, a
populációdinamika és a kvantitatív genetika fogalmainak segítségével.
A klasszikus megközelítésekkel összhangban hangsúlyozzuk, hogy a
természetes szelekció darwini fogalma szorosan összefügg a populációk
növekedésének szabályozottságával. Próbálkozásunkkal nem állunk
egyedül. A tudományfilozófus Gregory J. Cooper ökológiáról szóló
könyve a következő címmel jelent meg 2003-ban: A létért folyó
küzdelem tudománya. Az ökológia alapjairól. Ezzel a definícióval
összhangban azt állítjuk, hogy az ökológia mint „az élőlény-populációk
és társulások elterjedésének és számosságának kutatása” sikeresen
alapozható a létért való küzdelem és a fajképződés darwini
elméletére, ha hűen követjük Charles Darwin logikáját, és a
rátermettség fogalmát ennek megfelelően definiáljuk. Az ily módon
létrejött ökológiai elméletnek hét olyan – logikailag összefüggő –
alapelv a pillére, melyek génekre, klónokra és az ivarosan szaporodó
fajok populációira egyaránt érvényesek.
Bevezetés
Az ökológia egzakt
természettudományként történő felfogása és képviselete napjainkban is
komoly kihívást jelent. A természetrajzos szemlélet számos képviselője
a terepen szerzett, sokszor látszólag egymásnak ellentmondó
tapasztalatai alapján, illetve logikailag kikezdhetetlen elméleti
támpontok híján még ma is azt hangoztatja, hogy az ökológia – vizsgált
objektumainak egyedisége, bonyolultsága és a külső
feltételek változékonysága miatt – lényegileg különbözik
például a fizikától, és a hozzá hasonló, szigorúan elméleti alapokon
nyugvó tudományterületektől. Az ökológia helyzetének, eszközeinek,
lehetőségeinek megítélésében még a vezető elméleti ökológusok sem
teljesen egységesek, bár többségükben azért egy univerzális,
logikailag összefüggő alaptörvényekre épülő ökológia fejlesztésén
dolgoznak. A hetvenes, nyolcvanas évektől azonban az elmélettel
szemben megnyilvánuló szkepticizmus, mely szerint az egyes matematikai
modellek csupán egy vagy legfeljebb néhány, egymástól izolált
ökológiai probléma megoldásának specifikus eszközei, uralkodó
attitűddé vált az elméleti ökológián kívül. Bár az elméleti ökológia
művelői között jellemzően továbbra is kisebbségben vannak a
pragmatikusnak nevezhető kutatók, akik nem látnak lehetőséget
arra, hogy az egyedi ökológiai szituációkra kidolgozott
szimulációkat, modelleket a természetben általánosan érvényes
alapelvekhez és egységes matematikai elmélethez kössék, az egységes és
univerzális elmélet által irányított terepi, illetve kísérletes
kutatásokba vetett hit az ökológusok többsége számára elveszett. Ebben
a helyzetben viszont félő, hogy az ad hoc ötletek motiválta
elméleti és empirikus eredmények összefüggéstelen tömege maga alá
temeti az áttekintést, általános megértést igénylő szemléletet,
illetve az ezt képviselő kutatókat, oktatókat és diákjaikat egyaránt.
Az ökológia e „nyomorúságával”
szemben az evolúciós paradigmának óriási egységesítő ereje van
általában a biológiában, és különösen azon belül az ökológiában.
Darwin elmélete a létért való küzdelemről nemcsak az ún. „szintetikus
evolúcióelmélet” (Vida, 1981–85) alapja, hanem számos olyan
meghatározó jelentőségű ökológus munkássága is Darwin elméletén
alapult, mint Gause-é, akit a Lotka-Volterra kompetíciós modell
illusztrálására papucsállatkákon végzett szisztematikus laboratóriumi
vizsgálatsorozata, vagy Charles Eltoné, akit a táplálékpiramisai
tettek közismertté. Darwini alapelveken építkezett Hutchinson is, aki
a niche-elméletet újrafogalmazta, és széles körben népszerűsítette.
Egy sor nagyszerű kutató teremtett ebben a szellemben ma is élő
hagyományokat: David Lack (1947) a madárökológiában, John Harper
(1967) és David Tilman (1988) a növényökológiában, Ilkka Hanski (1999)
a (lepke-) populáció-hálózatok kutatásában, vagy Peter Turchin (2003)
az erdei rovarkártevők tömegesség-ingadozásaiból kiindulva a kaotikus
populációdinamika kutatásában. A század legnagyobb hatású elméleti
ökológusainak munkáját szintén Darwin motiválta, az általános
kompetíciós elméletet kidolgozó Alfred Lotkától az evolúciós ökológiát
elméletileg megalapozó Robert MacArthuron keresztül a strukturált
populációk általános evolúcióelméletének alapelveit formalizáló Hans
Metzig vagy a kompetíció-elméletet a fluktuáló környezetben élő
populációkra általánosító Peter Chessonig. A hetvenes évekre
kialakult, s azóta az ökológia integráns részévé vált az evolúciós
ökológia. Ez főként három, az evolúciós gondolat ökológiai
integrálására és különösen a populációgenetika és az ökológia
egységesítésére törekvő amerikai kutatónak köszönhető: a fizikus
képzettségű, fiatalon elhunyt Robert MacArthurnak, az evolúciógenetika
élő klasszikusának, Richard Lewontinnak és a mindkettőjükkel
klasszikus cikkeket publikáló, szintén fizikusnak tanult Richard
Levins filozófiai forrásból táplálkozó erőfeszítéseinek. Jelentős
szerepet játszott az ökológiai szempontból fontos, a fajok
elterjedését és tömegességét meghatározó tulajdonságok adaptációkként
történő felfogása is. Az adaptációkutatás technikai és fogalmi
nehézségeinek okán azonban mindeddig még a
populációbiológia sem kapott egységes elméletet (Lewontin, 2004), a
társulásökológiáról nem is beszélve (Chase – Leibold, 2003).
A darwini szelekciós elméletet, a
mendeli öröklődés-elméletet és a makroevolúciós elméletet
összeegyeztető modern szintézis masszív logikai problémája, ami
minden további integrálást ellehetetlenített, a természetes szelekció
és a diverzitás fogalmi és formális összekapcsolásának hiánya volt. Ez
a probléma Darwin számára még nem létezett. A fajképződést a
változatok folyamatos és fokozatos távolodásaként képzelte el, s
Robert Malthusra alapozó elméletéből az következett, hogy a speciációt
éppen a divergenciára történő természetes szelekció eredményezi. Az
adaptív tájkép igen vonzó metaforája, amit Sewal Wright, az elméleti
populációgenetika s egyben a modern szintézis egyik megalapozója már
1932-ben bevezetett, s nyomában George Gaylord Simpson a morfológiai
evolúció integrálója széles körben népszerűsített és terjesztett, maga
alá temette Darwin divergencia-elvét. Az adaptív tájkép metaforában a
természetes szelekció örökös hegymászással analóg egy olyan tájban,
amit a tulajdonságkombinációk adott, állandó környezetben érvényes
rátermettsége alakít ki (1. ábra). A legelőnyösebb
tulajdonságkombinációk a hegycsúcsok. Egy csúcsról csak úgy lehet
elkerülni, ha valamilyen véletlen folytán lesodródunk egy másik hegy
lábához. Az evolúció számára sorsdöntő véletleneket az elkóborló
egyedek által alapított új populációknak a szülőpopulációétól esetleg
lényegesen eltérő genetikai összetétele jelenti. Amíg az adaptív
domborzatot eleve adottnak tekintjük, amit csak külső körülmények
határoznak meg, s amit magának a szelekciónak a folyamata nem alakít
folyamatosan, addig a versengés csökkentése által hajtott divergencia
darwini koncepciója e képben nem vizualizálható. A divergencia-elv
háttérbe szorulásával párhuzamosan a kutatók a populációk genetikai
homogenizálódását kezdték szelekciós alaptendenciának tekinteni, s
éppen a genetikai polimorfizmus, azaz a fajokon, populációkon belüli
öröklődő változatosság fennmaradása igényelt speciális magyarázatot. E
szerint az öröklődő sokféleséget speciális genetikai mechanizmusok
tartják fenn, amelyeknek semmi közük a versengés csökkentéséhez: a
változatok többsége a szelekció szempontjából semleges és átmeneti,
vagy az allélok közti olyan gyakoriságfüggő szelekció tartja fenn,
mint amilyet például a hemoglobin gén egy változatára heterozigóta
egyedek előnye hoz létre a malária sújtotta területeken.
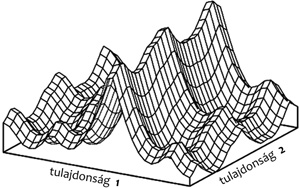
1. ábra • A Simpson–Wright-féle adaptív
tájkép: a tulajdonságok függvényében a rátermettség látható
Az integráció fogalmi nehézségeit
jelzi, hogy míg „a legrátermettebb fennmaradása” a szaporodási
egységet képező mendeli populációkon belül vitathatatlan alapelv ma
is, addig a kompetitív kizárás gondolata, azaz az egymáshoz hasonló,
versengő fajok populációi közötti természetes szelekció problémássá
vált. Hasonlóképpen, míg a faj rátermettségének fogalmát elfogadják,
és ma is használják az elméleti ökológiában, addig a
populációgenetikai rátermettség fogalmának összekötése a malthusi
paraméterrel, azaz a populációk növekedési ütemével, még a jövő
feladata egyes populációgenetikai monográfiák szerint is. Úgy véljük,
ezek a fogalmi problémák a formalizált elméletek alább tárgyalt
területein végbement nagymértékű fejlődés következtében mára
kezelhetőkké váltak. Bár a strukturált populációk elmélete (Metz et al.,
1992), a populációdinamika (Berryman – Kindlmann, 2008), a
társulásökológia (Chase – Leibold, 2003; Meszéna et al., 2006), a
kvantitatív genetika (Lynch – Walsh, 1997) és az adaptív dinamika (Meszéna
et al., 2005) mindegyike saját feltevés- és modellrendszert épített
ki, a rátermettség közös fogalma révén mégis összekapcsolhatók.
Darwin szelekciós elméletének
következetes alkalmazása fontos lépés lehet afelé, hogy az ökológia
diszciplináris kríziséből kilábaljunk. Első lépésként kurrens
ökológiai fogalmak használatával aktualizáljuk Darwin szelekciós
elméletét.
Darwin logikája
Darwin szelekciós elméletének
logikáját nemrégiben két amerikai kutató rekonstruálta a Nature-ben
megjelent évfordulós cikkükben. David Reznick és Robert Ricklefs
(2009) öt alapelve a következő: 1. az öröklődő változatosság, 2. az
utódtöbblet-termelés, 3. a populációnövekedési korlátok jelenléte,
amik a létért folyó küzdelemhez vezetnek, 4. a természetes szelekció:
a létért folyó küzdelemben szükségképpen rengeteg egyed pusztul el,
vagy marad utód nélkül, ami „a kedvező egyedi különbségek és
változatok megőrzéséhez és a károsak eltűnéséhez” vezet, és 5. az
adaptív divergencia: e küzdelemben azoknak a változatoknak a sikere
várható, amelyek „leginkább különböznek a közeli rokonaiktól a
túléléshez és szaporodáshoz kapcsolódó szükségleteikben”.
Mi Darwinnak e princípiumokhoz
kapcsolódó eredeti szövegeit ökológiai szempontból tekintjük át.
Darwin egyik kiindulópontja az volt, hogy a születések és a
halálozások számának ki kell egyenlítenie egymást ahhoz, hogy egy-egy
faj fennmaradjon, s ez az egyensúly a „növekedési korlátok” eredménye.
E korlátok megdöbbentően teljes listáját adja A fajok eredete
harmadik fejezetében, példái közt szerepelnek biotikus
kölcsönhatások (ragadozó–préda, gazda–parazita, növény–növényevő,
növény–beporzó szervezet) amik szabályozhatják a populációk
növekedését, és az éghajlat közvetett (a táplálék vagy a
megfelelő élőhely megváltozott mennyiségén keresztül realizálódó),
illetve közvetlen hatásai. Darwin hangsúlyozta, hogy utóbbiak közül a
biotikus kapcsolatokon keresztüli közvetett hatások az általánosak,
míg például a hideggel vagy az aszállyal vívott küzdelem csak „a fajok
földrajzi elterjedési területének határain zajlik”. Az öröklődő
változatok közti természetes szelekció abból a tapasztalati tényből,
illetve logikai szükségszerűségből következik, hogy a populációk
növekedését előbb vagy utóbb valamilyen tényező korlátozza. Darwin
szerint a természet az élőlény-populációkat jellemző exponenciális
növekedési képesség következtében telített. „A magas geometriai
növekedési ráta következtében minden terület teljesen tömve van.”
Bármely aprócska előny a versenytárs változattal szemben azonnal a
győztes változat egyedszámának növekedésében és a vesztes számának
csökkenésében nyilvánul meg. Így a létért folyó küzdelemben a jobb
változat/jobb faj fennmarad, míg a rosszabb kihal, s annál szorosabb
lesz a verseny, minnél hasonlóbb változatok között folyik: „A küzdelem
csaknem mindig a fajtársak közt lesz a legkeményebb, mivel azonos
körzetekben fordulnak elő, ugyanazt a táplálékot igénylik, és
ugyanazoknak a veszélyeknek vannak kitéve.” – azaz azok között a
változatok között, amelyeknek azonosak a növekedési korlátaik.
„Egy-egy faj változatai között csaknem ugyanilyen kemény lesz a
küzdelem, és néha megfigyelhetjük, hogy rövid idő alatt el is dől.” Ez
a gondolat, tehát a populációkon belüli természetes szelekció és a
populációk közti kompetitív kizárás közötti elvi hasonlóság veszett el
hosszú időkre, amikor Gregor Mendel örökléselméletét integrálták az
evolúciós elméletbe. Darwin ökológiai szempontból világosan és
konkrétan fogalmazott, amikor azt magyarázta, hogy a változatoknak
hogyan kell különbözniük ahhoz, hogy egymás mellett fennmaradhassanak:
„minnél különbözőbbé válnak egy faj leszármazottai felépítésben,
megjelenésben és szokásokban, annál inkább képesek lesznek arra, hogy
a természet államának sok és szélesen változatos pozícióit betöltsék,
s így egyedszámukat növeljék”. Még konkrétabban fogalmazott, amikor
egy „elképzelt példával” illusztrálta mondandóját: „Vegyük a farkas
esetét, amely sokféle vadra vadászik. Egy részüket az erejével, más
részüket az ügyességével, megint másokat a gyorsaságával ejti el.
Tételezzük fel, hogy a leggyorsabb zsákmányának, mondjuk egy
szarvasnak, az adott vidéken végbement valamiféle változás miatt
megnövekedett a létszáma, a többi prédaállat száma pedig lecsökkent
abban az évszakban, amikor a farkast a leginkább szorítja az
élelemszerzés. Ilyen körülmények között a legfürgébb és a legsoványabb
farkasoknak lesz a legjobb esélyük a túlélésre, és így a fennmaradásra
és kiválasztódásra. […] a Catskill-hegységben a farkasnak két
változata él: egy agár alkatú, amely szarvasra vadászik, és egy másik,
tömzsibb, rövidebb lábú, amely gyakrabban támadja meg a juhászok
nyájait.” Darwin dinamikus gondolkodásmódja itt szépen megnyilvánul: a
kétféle farkaspopuláció (a szarvason, illetve a juhon élő) növekedési
korlátai különbözőek, ami mindkettő számára lehetővé teszi, hogy
növeljék a számosságukat, azaz, hogy együtt éljenek. Ez a fajképződés
alapja. Figyeljük meg, hogy ez a divergencia-elv feltételezi, hogy a
farkasok között nem jelenhetnek meg olyan variánsok, amelyek egyaránt
jók a juhok és a szarvasok elejtésében (lásd: kényszerek).
A dinamikus gondolkodásmódon túl
Darwin tisztában volt a változások véletlenszerű természetével is: „A
puszta véletlen, nevezhetjük így is, elérheti, hogy egy változat
valamilyen tulajdonságában különbözzön a szüleitől, majd e változat
utódai szintén ugyanebben a tulajdonságukban kissé nagyobb mértékben
különbözzenek tőlük, de ez önmagában soha sem fogja megmagyarázni
azokat a nagymértékű különbségeket, amelyeket egy nemzetség különböző
fajai közt általában találunk.” Darwin tárgyalta a semleges változatok
lehetséges sorsát is: „A se nem káros, se nem előnyös változatokat a
természetes szelekció nem érinti, és vagy fluktuáló elemek maradnak –
ahogy azt talán néhány polimorf fajnál látjuk –, vagy végül is
fixálódnak a szervezet és a feltételek természetétől függően.”
Darwin gondolkodásmódjának ökológiai logikája a fajok exponenciális
növekedési kapacitásának és az öröklődő változatok keletkezésének
felismerésével indul, a növekedési korlátok elkerülhetetlenségének
tényével folytatódik, majd az e korlátok divergenciájára állandóan
folyó természetes szelekció gondolatával zárul. Ez a logika bizonyára
nyilvánvaló volt az olyan kiváló ökológusok számára, mint Elton vagy
David Lack, akik explicit módon összekapcsolták a természetes
szelekció erősségét a populációk dinamikájával, illetve a
növekedésszabályozást a jellegeltolódással
|
|
|
és a fajképződéssel. Hutchinson a niche fogalmát a Gause-féle
kompetitív kizárási elvvel kapcsolta össze, míg MacArthur Ronald
Fisher szelekciós alaptételét általánosította ugyanebben a szellemben
a szabályozott növekedésű populációkra. Mindezek ellenére Darwin
robusztus gondolatai elsüllyedtek a technikai nehézségek mocsarában,
mind az ökológiában, mind az evolúcióelméletben.
Darwin logikájának aktualizálása
Szaporodási egységek
Modern értelmezésünk kiindulópontja három
szaporodási vagy reprodukálódó egység, típus elkülönítése. Bármilyen
reprodukálódó egység populációja képes exponenciális növekedésre. A
reprodukálódó egységek lehetnek egy fajhoz tartozó, ivarosan szaporodó
egyedek (populációjuk a mendeli populáció), ivartalanul szaporodó
egyedek, vagyis klónok (populációjuk a klónpopuláció) valamint az egy
genetikai lokuszhoz tartozó génváltozatok, azaz allélok (egy allél
másolatainak populációja). Akkor van e szaporodási egységek öröklődő
változatai között szelekció, ha azok populációinak tömegessége
különböző ütemben növekszik: a legnagyobb növekedési rátájú változat
exponenciálisan túlnövi az alacsonyabb ütemben szaporodó változatokat.
A különböző tulajdonsággal rendelkező egyedek rátermettsége
„annak a populációnak a növekedési rátája, ami kizárólag az adott
tulajdonságú egyedekből áll.” Ez a definíció mindhárom szaporodási
egység populációjára kiterjeszthető. Az ivarosan szaporodó egyedek
esetén a reprodukálódó tulajdonság a faji hovatartozás. A rátermettség
az esetek többségében a külső körülmények és a többi jelenlévő
változat tömegességének is függvénye.
Az ivarosan szaporodó diploid szervezetek
populációiban nem mindig a legrátermettebb genotípusú egyedek
„győznek”: gondoljunk arra, hogy ha a heterozigóták rátermettsége a
legmagasabb – például a maláriával fertőzött területeken a hemoglobin
sarlósejtes vérszegénységet okozó változatára heterozigóta emberek
rátermettsége magasabb, mint a csak a normális vagy csak a mutáns
allélt hordozó homozigótáké – akkor sem lesz a népességben mindenki
heterozigóta. Az öröklés módja miatt a heterozigóták utódai között
mindig előfordulnak homozigóták. Az egyedek által hordozott
allélkombinációk minden ivarsejtképződéskor felbomlanak, így nem
reprodukálódnak, azaz a genotípus nem szaporodási egység. Maguk az
allélok másolódnak, s másolataik száma mind szaporodás előtt, mind
utána megszámlálható, azaz a darwini rátermettség-fogalom allélokra
alkalmazható, míg ugyanez a genotípusokról, az allélok egy-egy
egyedben előforduló kombinációiról nem mondható el. Mivel ugyanazon az
alapon beszélhetünk allélok, klónok és mendeli populációk
rátermettségéről, az allélok közti természetes szelekció folyamata
analóg a klónok vagy a fajok mendeli populációi közti ökológiai
kompetíció folyamatával. Így felfogva a „kompetitív kizárás elve” és
„a legrátermettebb fennmaradása” felcserélhető kifejezések.
Populációreguláció és a létért folyó küzdelem
Amikor Darwin a „létért folyó küzdelem” fogalmát
bevezette, megjegyezte, hogy „általánosságban, metaforikus értelemben”
használja, azaz nem tudta a megfelelő fogalmat absztrakt szinten
definiálni. A fajok eredete szövegére és Darwin fent vázolt
logikájára támaszkodva úgy gondoljuk, hogy a „létért folyó küzdelem”
néhány kivételtől eltekintve a szabályozó tényezők által indukált
versengéssel azonos. A növekedési korlátok ugyanis rendszerint
szabályozó tényezők (táplálék, ragadozó, parazita, tér stb.) vagy
esetenként olyan extrém külső feltételek, amelyek között az egyedek
puszta fennmaradásukért küzdenek. A modern evolúciós elméletekkel
összhangban ezt a kétfajta küzdelmet kétféle szelekciós folyamatnak
feleltethetjük meg. A „kihalással történő természetes szelekció” az
egyedek elemekkel folytatott harcából következik, és mind korlátlanul
növekvő, mind szabályozott növekedésű populációkban bekövetkezik. Ez
esetben a kevéssé alkalmas változatok elpusztulnak, mert populációjuk
növekedési kapacitása negatív az adott körülmények között. Például a
letális vagy terméketlenséget okozó mutációk minden körülmények között
kihalnak, azaz a többi változat jelenléte nem befolyásolja sorsukat. A
„kizárással történő természetes szelekció” esetén mindegyik változat
növekedési kapacitása pozitív, azaz a tisztán egy-egy változat
egyedeiből álló populációk növekednének. Ez esetben korlátozó tényezők
hiányában az összes változat fennmaradhatna, bár arányuk
exponenciálisan változna. Ezért „kizárással történő természetes
szelekció” nem fordulhat elő exponenciálisan növekvő populációkban,
csak szabályozott növekedésűekben.
Fontos különbséget tenni a növekedési kapacitás
(r0) – ami a változat populációjának növekedését
akkor jellemzi, amikor a populációdenzitás kicsi (azaz a növekedés
korlátlan) és nincsenek jelen más változatok –, valamint a
rátermettség mértéke, a pillanatnyi (fejenkénti) növekedési ráta (r)
között, ami a körülmények (biotikus és abiotikus) és a
populációdenzitás függvényében változik. Ha egy szabályozott
növekedésű populációban egyetlen legjobb változat van, annak
egyensúlyban nulla lesz az átlagos pillanatnyi növekedési rátája, míg
a többi, kiszoruló változaté negatív.
Kizárás és robusztus együttélés
Implicit darwini gondolat az, hogy egy-egy változat fennmaradása
vagy kizáródása azon múlik, hogy a versengő változatok egyedszáma
hogyan szabályozódik. Idézzük fel a farkas példáját! Ha a két változat
egyedszámát különböző táplálék szabályozza (juh, illetve szarvas),
akkor a természetes szelekció egyes jellegek divergenciájához, végső
soron fajképződéshez vezet. Ez a biológiai sokféleség gyökere. Darwin
gondolata összekapcsolható továbbá a gyakoriságfüggő szelekció modern
fogalmával, a növekedésszabályozásért felelős visszacsatolás
dimenziószámával és a robusztus együttélés feltételeivel.
Gyakoriságfüggő szelekcióról akkor beszélünk, amikor az egyes
változatok rátermettsége, azaz populációik növekedési rátája függ
attól, hogy milyen egyéb változatok milyen gyakorisággal vannak jelen.
Ha a rátermettség csak a populációk összlétszámától függ, akkor
denzitásfüggő szelekcióról beszélünk, és az ún. „visszacsatoló
környezet” egydimenziós. Ilyenkor a változatok közti versengésnek
egyetlen győztese van, amelyet valamelyik extrémum szabály jósol meg:
például forrásért folyó versengés esetén az R*-szabály (a
legalacsonyabb tápanyag-koncentráción megélő növényfaj [allél, klón]
győz) érvényes. Együttélés csak akkor lehetséges, ha a környezeti
visszacsatolás több, mint egy dimenziós. Gyakoriságfüggő szelekció
esetén a ritka változat előnye stabil együttéléshez vezet. Az
együttélés csak akkor lesz robusztus, azaz ellenálló a környezeti
változásokkal szemben, ha az együttélő populációkat más-más tényezők
szabályozzák, vagy közös szabályozó tényezők esetén a szabályozás
módjában nagymértékben különböznek. Mind a szabályozó tényezőkre
kifejtett hatás mértékében, mind a szabályozó tényezőkre való
érzékenység mértékében jelentősen különbözniük kell az egyes
változatoknak ahhoz, hogy a környezeti tényezők széles tartományában
együtt éljenek.
Kényszerek és csereviszonyok
Az evolúciós ökológiában szokás „darwini
démonnak” nevezni az olyan elképzelt élőlényt, amely egyszerre
végtelen hosszú életű és végtelenül szapora. Ha szorosan követjük
Darwin logikáját, akkor ez a „darwininak” nevezett „szuperorganizmus”,
egyáltalán nem darwini. Darwin egyensúlyi szemlélete szerint ugyanis a
növekedési korlátok eleve lehetetlenné teszik egy ilyen démon
létezését. Azaz egy halhatatlan, folyamatosan szaporodó gép helyett az
igazi darwini démon olyan élőlény, amely maximális hatékonysággal
fogyaszt minden forrást, foglal el minden teret, menekül el minden
ragadozó elől, kerül el minden betegséget. Ez a változat minden más
változatot kiszorítana, s egyeduralkodóként végül egyensúlyban élne
abiotikus szabályozó tényezőivel. Azaz az egyedi szerveződés
csereviszonyokhoz vezető belső kényszerei nélkül nem lenne lehetséges
a fajképződés.
Egy másik nem triviális következménye Darwin
divergencia-elvének, hogy minden populációra a szabályozó tényezők
egyedi kombinációja hat, és szelektál a lehetséges változatok között.
Egy-egy fajnak számos, egymástól genetikailag eltérő populációja
létezik változatos környezeti feltételek között, amelyek különböző
szintű halálozáshoz és termékenységhez vezetnek. A populációnövekedés
szabályozottságának következtében néhány életmenet-tulajdonság
(termékenységi ráták, halálozási ráták, ivarérés kora, reproduktív
élethossz stb.) vagy denzitás- vagy gyakoriságfüggő kell hogy legyen.
Ezért a nem szabályozó környezeti feltételek változása nemcsak
közvetlenül szelektálhat a változatok között, hanem közvetve, az egyes
változatok denzitásának megváltoztatásán keresztül is, az
életmenet-tulajdonságok denzitásfüggésének következtében. Az
életmenet-jellemzők közti csereviszonyok és a növekedésszabályozás
közös eredményeként sokszor megfigyelhető, hogy az optimális
életmenet-stratégiák az egyensúlyi populációdenzitás függvényében
változnak. Ezt a mintázatot az r/K-szelekció elmélete tette
közismertté.
Fajképződés
A fajok eredetének – a Darwin számára legfontosabb megoldandó
problémának – darwini magyarázata, mely a divergens evolúciót a
kompetíciós nyomás csökkentése felé ható szelekcióval hozza közvetlen
kapcsolatba, a mai elméleti megközelítések közül a
kompetitív/adaptív fajképződés elméletével mutat nyilvánvaló
rokonságot. A speciáció e darwini magyarázata a szimpatrikus (térbeli
izoláció nélküli) mechanizmusok jelentőségét hangsúlyozza, ami éles
ellentétben áll Ernst Mayr (2001) jellemzően allopatrikus
szemléletével, aki számára ez egyértelműen alárendelt jelentőségű.
Darwin számára a divergenciára történő szelekció a domináns fajképző
mechanizmus, a különböző élőhelyekhez való alkalmazkodás a versengés
enyhítésének csak egy speciális módja. A modern, formalizált
niche-elmélet, az adaptív dinamika és az adaptív fajképződés elméletei
a korábban tárgyalt értelemben egyértelműen a darwini speciáció
elveire és a szimpatrikus fajképződés gyarapodó terepi bizonyítékaira
épülnek. Ennek megfelelően a darwini speciáció szemléletmódja szinte
változatlan tartalommal, de modern elméleti és empirikus módszerek
birtokában, velük összhangban szolgálja a fajok eredetének és
együttélésének megértését.
Az ökológia darwini alapelvei
Meggyőződésünk, hogy néhány, a darwini gondolat
magvát képező alapelv következetes szem előtt tartásával az ökológia
legfontosabb problémái logikailag egységes szemléleti keretben
tárgyalhatók. Az alább felsorolt hét alapelvet az ökológia
„kristályosodási gócainak” tekintjük, melyek köré megfelelően
strukturált, koherens és a gyakorlatban is alkalmazható elméleti tudás
szervezhető. Alább csak az alapelvek felsorolására szorítkozunk –
részletesebb kifejtésükre, logikai és módszerelméleti kapcsolataik
elemzésére készülő tankönyvünkben (Pásztor et al., in prep.)
kerül sor.
• Exponenciális növekedési kapacitás: Szabályozó
visszacsatolások hiányában bármely önreprodukáló egységekből
álló populáció létszáma exponenciálisan növekszik.
• Öröklődő változatosság: Minden
önreprodukáló egységekből álló populációban előfordulnak másolódási
hibák, amelyek az utódokban öröklődnek, és befolyásolhatják azok
szaporodási/túlélési sikerét.
• Szabályozott növekedés: Minden növekvő
populáció előbb-utóbb a saját további növekedésének gátjává válik: a
populációnövekedés negatív visszacsatolással szabályozott folyamat,
mely végül a populáció nulla gyarapodását eredményezi hosszú távon.
• Kompetitív kizárás: Ha több populáció
növekedését egyetlen környezeti tényező szabályozza, akkor
mindig csak az az egy populáció marad fenn (nulla növekedési
ütemmel), amelyet ez a tényező a legkevésbé gátol a növekedésben; a
többi populáció kihal.
• Robusztus együttélés: Különböző önreprodukáló
egységek populációinak tartós együttélése kizárólag akkor
lehetséges, ha populációnövekedésük szabályozásában elegendő
mértékben eltérnek egymástól.
• Csereviszonykorlátok: Az egyedi tulajdonságok
változatossága egyedszerveződési vagy energetikai okokból mindig
korlátozott; a rátermettség komponensei egymással többnyire
csereviszonyban állnak.
• Sztochaszticitás: A populációk véges méretéből
eleve következik az ökológiai jelenségek sztochaszticitása. Az
egyedek véges számából következően változataik száma is véges, és
bármelyik változat pusztán véletlenül is kihalhat.
A fenti alapelvek univerzálisak,
amennyiben minden önreprodukáló egység populációira szükségszerűen és
általánosan érvényesek, másrészt koherensek abban az
értelemben, hogy levezethető belőlük egy, a szabályozott
populációnövekedés elvén alapuló, integrált és formalizált ökológiai
elmélet. A fajok eredete központi fogalma, a „létért folyó
küzdelem” ebben a felfogásban a szabályozott populációnövekedés
metaforája, amely a fenti hét alapelv kontextusában egyszerre adhat
magyarázatot a természetben annyiszor megfigyelt, de megnyugtatóan
máig meg nem értett biológiai sokféleség evolúciós létrejöttére és az
azt fenntartó ökológiai mechanizmusok működésmódjára.
Kulcsszavak:
Darwin, létért folyó küzdelem, populációreguláció, ökológia, evolúció,
rátermettség, elmélet, alapelvek
IRODALOM
Berryman, Alan A. – Kindlmann, Pavel (2008):
Population Systems: A General Introduction. 2. ed. Springer
Chase, Jonathan M. – Leibold, Matthew A. (2003):
Ecological Niches, Linking Classical and Contemporary Approaches.
The University of Chicago Press, Chicago
Cooper, Gregory J. (2003): The Science of the
Struggle for Existence: On the Foundations of Ecology. Cambridge
University Press, Cambridge
Darwin, Charles (1872): The Origin of Species
by Means of Natural Selection, Or the Preservation of Favoured Races
in the Struggle for Life. 6th ed. John Murray, London,
http://darwin-online.org.uk/
Hanski, Ilkka (1999): Metapopulation Ecology.
Oxford University Press, New York
Harper, John L. (1967): A Darwinian Approach to
Plant Ecology. Journal of Ecology. 55, 247–270.
Lack, David (1947): Darwin’s Finches.
Cambridge University Press, Cambridge
Lewontin, Richard C. (2004): Building a Science
of Population Biology. In: Singh, Rama Shankar – Uyenoyama, Marcy K. (eds.):
The Evolution of Population Biology. Cambridge University Press,
Cambridge, 7–20.
Lynch, Michael – Walsh, Bruce (1997):
Genetics and Analysis of Quantitative Traits. Sinauer Associates,
Sunderland, MA.
Mayr, Ernst (2001): What Evolution is.
Basic Books, New York
Meszéna Géza – Gyllenberg, M. – Jacobs, F. J. –
Metz, J. A. J. (2005): Link between Population Dynamics and Dynamics
of Darwinian Evolution. Physical Review Letters. 95, 078105.
Meszéna Géza – Gyllenberg, M. – Pásztor L. –
Metz, J. A. J. (2006): Competitive Exclusion and Limiting Similarity:
A Unified Theory. Theoretical Population Biology. 69,
68–87.
Metz, Hans A. J. – Nisbet, R. M. – Geritz, S. A.
H. (1992): How Should We Define “Fitness” for General Ecological
Scenarios? Trends in Ecology & Evolution. 7,
198–202.
Pásztor Erzsébet – Botta-Dukát Z. – Czárán T. –
Magyar G. – Meszéna G. (in prep.): Theory Based Ecology. A
Darwinian Approach. Oxford University Press
Reznick, David N. – Ricklefs, Robert E. (2009):
Darwin’s Bridge between Microevolution and Macroevolution. Nature.
457, 837–842.
Tilman, David (1988): Plant Strategies and
the Dynamics and Structure of Plant Communities. Princeton
University Press, Princeton
Turchin, Peter (2003): Complex Population
Dynamics: A Theoretical/Empirical Synthesis. Princeton University
Press
Vida Gábor (szerk.) (1981–85): Evolúció
I–V. Natura, Budapest
1 A cikk az OTKA K67907 és K49689 számú kutatási
projektjeinek támogatásával készült.
<
|
|
