|
|
Az élő szervezetek működésében meghatározó szerepet
játszanak a sejtekben található dinamikus fehérjerendszerek, amelyek
azok szerkezeti és funkcionális vázát biztosítják. Ennek egyik
nélkülözhetetlen eleme az aktin sejtváz vagy aktin citoszkeleton. Az
aktin citoszkeleton esszenciális szerepet játszik szinte minden
sejtfolyamatban. Meghatározó a változatos sejtfunkciók
kialakításában, azok pontos térbeli és időbeli szabályozásában és
összehangolásában. Az aktin sejtváz alapvető alkotóeleme az
aktinfehérje. Az aktin a sejtben két formában fordulhat elő;
globuláris monomerként vagy az ezekből kialakuló szálszerű
filamentumként (Bugyi – Carlier, 2010). A monomerek spontán módon is
filamentumokba szerveződhetnek, ezt polimerizációnak nevezzük. A
folyamat első szakasza, a nukleáció során két, ill. három monomer
összekapcsolódása révén nukleációs magocskák alakulnak ki. Ezek
további monomerek hozzákapcsolódása következtében növekednek
(elongáció), így létrejönnek a filamentumok. A spontán polimerizáció
viszonylag lassú folyamat a nukleációs magok instabilitása miatt,
így egy filamentum kialakulása akár órákat is igénybe vehet.
Gondoljuk meg, hogy ez a sejt működése szempontjából nem szerencsés,
egyrészt a sejt „óráját” tekintve rendkívül lassú, másrészt térben
és időben szabályozatlan, anarchikus filamentum képződéshez vezetne.
Hogyan képes a sejt ezt a folyamatot felgyorsítani, azaz
katalizálni, valamint szabályozni? Miként tudnak ezek az „egyszerű”
aktinszálak változatos funkciókat létrehozni? Ezek a kérdések az
aktin szabályozó fehérjéinek színes világához vezetnek minket,
amelyek az aktin sejtváz működésének finomhangolását végzik.
A sejtben az aktin filamentumok polimerizációjának
szabályozásáért apró molekuláris fehérjegépezetek, az úgynevezett
összeszerelő faktorok felelősek (Bugyi – Carlier, 2010). Ezek közé
tartozik a formin fehérjecsalád is. A forminfehérjék széles
taxonómiai eloszlást mutatnak, az egysejtű élőlényektől az emberig
találjuk meg őket. Az 1990-es évek végi felfedezésük óta a család
számos taggal bővült, jelenleg tizenöt humán formint kódoló gént
ismerünk, amelyről átíródó fehérjék hét családba sorolhatóak.
Bizonyos forminok diszfunkciója betegségekhez vezethet, mint a
krónikus limfómás leukémia, petefészek-elégtelenség vagy különböző
típusú rákos megbetegedések. A forminok doménszerkezetű molekulák,
azaz egyes régióikhoz jól meghatározott funkciók köthetőek.
Azonosításuk az úgynevezett formin homológia (FH) 1 és 2 domének
megléte alapján történik. Ma már tudjuk, hogy az FH2 domén a fehérje
legkonzerváltabb egysége, amely alapvető fontosságú az aktinnal
kialakított kölcsönhatásban, az FH1 domén pedig egy másik aktinkötő
fehérjéhez, a profilinhez kapcsolódik. Hogyan működik ez a
fehérjegépezet, miként képes az aktin filamentumok összeszerelődését
elősegíteni?
Ezzel a kérdéssel a 2000-es évek elején, a Dia
(Diaphanous-related) formin család működésének szerkezeti és
működésbeli jellemzőinek vizsgálata révén kezdtem el foglalkozni.
Kutatásaink során partnerünkkel (Prof. Alfred Wittinghoffer, Max
Planck Intézet, Dortmund, Németország) elsőként határoztuk meg a Dia
formin izolált FH2 doménjének atomi felbontású térszerkezetét. A
funkcionális vizsgálataink azonban arra világítottak rá, hogy ez a
régió nem elegendő az aktin polimerizációjának gyorsításához, épp
ellenkezőleg, gátolja ezt a folyamatot. Azonosítottunk egy rövid (72
aminosavból álló), az FH2 domént megelőző szakaszt, ami két FH2
domén összekapcsolódását, vagyis dimerizációját biztosíthatja.
Megmutattuk azt is, hogy a dimerizáció alapvető szerepet játszik
abban, hogy a Dia fehérjék hatékony aktin filamentumösszeszerelő
faktorként működjenek, mégpedig feltételezhetően úgy, hogy az FH2
dimer az instabil nukleációs magok szerkezetét stabilizálja
(Shimada, 2004).
A későbbiekben kutatócsoportom érdeklődése a
DAAM-család (Dishevelled-associated activator of morphogenesis)
forminjaira terelődött. Bár a forminok FH2 régiója alapvető
fontosságú a forminok aktinnal kialakított kölcsönhatásában, érdekes
módon azt találtuk, hogy a DAAM ezen régiója önmagában semmilyen
módosulást nem okoz az aktin sejtvázban, míg az FH1–FH2 domének
együttesen drámai morfológiai és dinamikai változásokat képesek
előidézni (Barkó et al., 2010). A megfigyeléseink pontosabb
megértése érdekében izolált FH2 és FH1–FH2 doméneknek az aktin
polimerizációjára kifejtett hatásait tanulmányoztuk teljes belső
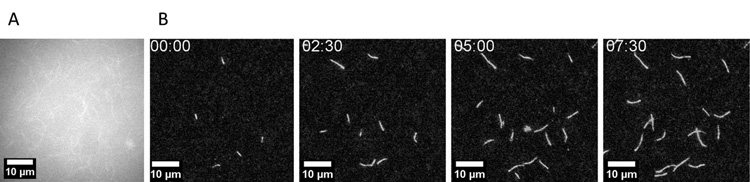
visszaverődésen alapuló fluoreszcencia-mikroszkópia (TIRFM)1
módszerével. A technika viszonylag új, első gyakorlati alkalmazása
az 1990-es évek végére tehető. Ez a speciális fénymikroszkópiai
eljárás egy optikai jelenség, a teljes belső visszaverődés során
keletkezett, ún. evaneszcens (tovatűnő) elektromágneses mező révén
lehetővé teszi a
|
|
|
vizsgálandó minta csupán egy vékony rétegének
szelektív megvilágítását. Így, a hagyományos
fluoreszcencia-mikroszkópiai technikákkal szemben a TIRFM
alkalmazása segítségével egyedi aktin filamentumokat tehetünk
láthatóvá (1. ábra). Ez pedig lehetőséget ad a polimerizáció
egyes szakaszaira (nukleáció és elongáció) kifejtett hatások
elkülönített vizsgálatára.
A módszer alkalmazásával azt találtuk, hogy mind az
FH1–FH2, mind pedig az FH2 domén jelenlétében lényegesen több aktin
filamentum volt megfigyelhető, ami arra utal, hogy ezek a régiók
képesek felgyorsítani a nukleációs fázist. Azonban, meglepő módon a
DAAM ezen szakaszai szinte teljes mértékben gátolták a filamentumok
növekedését, az elongációt. Ez a megfigyelés egy külső faktor
irányába terelte a figyelmünket. Más forminokkal kapcsolatos
vizsgálatok alapján már ismert volt, hogy az FH1 régió profilint
képes kötni. Így a fenti vizsgálatainkat elvégeztük ennek a
fehérjének a jelenlétében is. Igazán izgalmas megfigyeléseket
tehettünk; míg az FH2 domén továbbra is gátoló hatást fejtett ki, az
FH1–FH2 régió az eddigiekkel ellentétben profilin jelenlétében
elősegítette a filamentumok növekedését. Mindezek arra engedtek
következtetni, hogy a profilin molekuláris kapcsolóként szolgál a
DAAM funkciójában; a filamentumnövekedést gátló faktorból hatékony
filamentumösszeszerelő gépezetté alakítja (Barkó et al., 2010).
Kutatásainkkal egy időben más családból származó forminok izolált
FH1–FH2 doménjeinek aktivitásai is feltérképezésre kerültek. Az
átfogó vizsgálatoknak köszönhetően ismertté vált, hogy az FH1–FH2
szakaszok nélkülözhetetlenek a formin fehérjecsalád megfelelő
funkcionalitásában. Talán mondhatjuk, hogy ezek a felismerések
jelentették a forminkutatás első forradalmi időszakát.
De ne gondoljuk, hogy a forminfehérjék nem
tartogatnak több meglepetést. Együttműködő partnerünk (Mihály
József, MTA Szegedi Biológiai Kutatóközpont Genetikai Intézet)
Drosophila melanogaster (ecetmuslica) modell rendszerben végzett
vizsgálatai feltárták, hogy a DAAM-formin működése igen változatos
biológiai funkciókban érhető tetten, és nélkülözhetetlen a különböző
szervek morfogenezise, valamint megfelelő működése során. A DAAM, az
aktin sejtváz alapvető szabályozó molekulája például az idegsejtek
axonális nyúlványainak létrehozásában az idegrendszerben és az
idegsejtek közötti kapcsolatok kialakításában (Matusek et al.,
2008), a vázizom szarkomerikus vékony filamentumainak
összeszerelődésében és az izomösszehúzódásban (Molnár et al., 2014),
valamint a légzőrendszer funkcionalitásában (Matusek et al., 2006).
E megfigyelések izgalmas kérdéseket vethetnek fel. A DAAM-formin
polimerizációt elősegítő hatása hogyan képes ilyen változatos
funkciókat eredményezni? Ez az aktivitás áll-e a DAAM különböző
biológiai funkcióinak hátterében? Módosítják-e egyéb tényezők az
FH1–FH2 aktinnal kialakított kölcsönhatását az egyes funkciókhoz
való adaptációhoz? Amennyiben igen, mik ezek az elemek, és miként
működnek? Úgy gondolom, hogy ezek a felvetések a forminkutatás új és
izgalmas irányát indították el, amely minden bizonnyal érdekes
felfedezésekhez fog vezetni, amelyeknek köszönhetően még
részletgazdagabb képet kaphatunk majd erről a lenyűgöző molekuláris
gépezetről.
Kulcsszavak: aktin, formin, polimerizáció,
fluoreszcencia-mikroszkópia
IRODALOM
Barkó Szilvia – Bugyi Beáta et al. (2010):
Characterization of the Biochemical Properties and Biological
Function of the Formin Homology Domains of Drosophila DAAM. The
Journal Of Biological Chemistry. 285, 17, 13154–13169.
DOI: 10.1074/jbc.M109.093914
Bugyi Beáta – Carlier, Marie-France
(2010): Control of Actin Filament Treadmilling in Cell Motility.
Annual Review Of Biophysics. 39, 449–470. DOI:
10.1146/annurev-biophys-051309-103849
Matusek Tamás – Djiane, Alexandre et al.
(2006): The Drosophila Formin DAAM Regulates the Tracheal Cuticle
Pattern through Organizing the Actin Cytoskeleton. Development. 133,
5, 957–966. DOI: 10.1242/dev.02266 •
WEBCÍM
Matusek Tamás – Gombos Rita et al. (2008):
Formin Proteins of the DAAM Subfamily Play a Role during Axon
Growth. Journal of Neuroscience. 28, 49. DOI:
10.1523/JNEUROSCI.2727-08.200 •
WEBCÍM
Molnár Imre – Migh Ede et al. (2014): DAAM
Is Required for Thin Filament Formation and Sarcomerogenesis during
Muscle Development in Drosophila. PLOS Genetics. 10, 2. DOI:
10.1371/journal.pgen.1004166 •
WEBCÍM
Shimada, Atsushi – Nyitrai Miklós et al.
(2004): The Core FH2 Domain of Diaphanous-Related Formins Is an
Elongated Actin Binding Protein that Inhibits Polymerization.
Molecular Cell. 13, 4, 511–522. DOI: 10.1016/S1097-2765(04)00059-0 •
WEBCÍM
|
|