|
|
A mágneses rezonanciás képalkotás (MRI) a
röntgentechnológiák (például a számítógépes tomográfia) mellett az
orvostudomány jelenleg használt egyik legfontosabb diagnosztikai
eljárása. A módszer lényege, hogy az egyes atommagok képesek
abszorbeálni egy adott mágneses sugárzást, majd ezt a
többletenergiát rádióhullámú jelként adják le a mágneses tér
megszűnésekor. Megfelelő eljárással a test kemény, illetve
lágyszövetei láthatóvá tehetők, anatómiai pontosságú képet nyújtva a
test belső szerkezetéről.
Ez az eljárás azonban nemcsak strukturális és anatómiai vizsgálatok
elvégzésére alkalmas, hanem lehetővé teszi a különböző szövetek
dinamikus állapotváltozásainak nyomon követését is. A funkcionális
MRI (fMRI) ezt a lehetőséget használja ki a testen belüli
hemodinamikai folyamatok vizsgálatára, amelyekből következtethetünk
például az agyi aktivitás mintázatára. Az idegsejtek (neuronok)
működése igen energia- és oxigénigényes folyamat, és mivel ezek a
sejtek nem rendelkeznek saját cukortartalékokkal, a szervezet
fokozott vérellátással oldja meg a fogyasztás utánpótlását. A
vértöbblet egyúttal nagyobb arányú oxigenizált hemoglobint is
jelent, aminek a mennyiségét fMRI segítségével mérni tudjuk. Tehát
áttételesen a vér oxihemoglobin-tartalmának változását figyelve
azonosíthatók az agyban azok a területek, amelyek aktívabbak vagy
esetleg gátoltak, miközben az agy különböző feladatokat hajt végre
egy kísérlet közben. Így más módszerekhez képest az fMRI igen
részletes bepillantást enged az agy működésébe, különösen a működés
térbeli megnyilvánulását illetően. Az 1990-es évekbeli bevezetése
óta az fMRI a kognitív tudományok kedvelt eszközévé vált
(Logothetis, 2008), főleg azért, mert teljességgel nem invazív
eljárás, azaz a kísérleti alanyokat a vizsgálat során semmilyen
károsodás nem éri.
fMRI embereken és állatokon
Az emberi agy működésének, az ingerek feldolgozásának és a
pszichológiai folyamatok jobb megértésének elengedhetetlen
feltétele, hogy evolúciós perspektívába tudjuk helyezni a vizsgált
jelenségeket. Ehhez viszont értelemszerűen nem elegendőek a saját
fajunkkal végzett kísérletek, hanem közelebbi és távolabbi
rokonainkat is hasonló vizsgálatoknak kell alávetni. Egészen
mostanáig az ilyen összehasonlító neurobiológiai kutatások a
laborban tartott modellfajok, például rágcsálók (egerek, patkányok,
nyulak), illetve majmok (elsősorban makákók, selyemmajmok)
bevonásával történtek, mivel e fajok esetében különböző invazív
módszerek alkalmazásával már hatalmas anatómiai, illetve
idegélettani ismeret állt rendelkezésre. A laborállatokon végzett
fMRI-vizsgálatok azonban módszerükben lényegesen eltérnek az ember
esetében alkalmazott eljárástól. Az fMRI-vizsgálatokhoz
elengedhetetlen ugyanis, hogy az alany mozdulatlanul feküdjön,
miközben az agyat a különféle vizuális, illetve akusztikus ingerek
érik, hiszen csak ekkor lehet pontosan meghatározni az agyi
aktivitás helyét egy adott mágneses térhez képest. Laboratóriumi
állatok esetében a mozdulatlanságot különböző kényszerítő eszközök
alkalmazásával érik el (például bódítás vagy a test és a fej fizikai
rögzítése) (Van der Linden et al., 2007). Bár az ilyen neurobiológai
kutatások is sok izgalmas eredményt tártak fel, az alkalmazott
eljárások miatt kétséges az emberrel való közvetlen
összehasonlíthatóság. Túl messzire menő következtetéseket nem lehet
levonni a laboratóriumi állatokon végzett fMRI-vizsgálatok korlátai
és az elérhető fajok szűk köre miatt, illetve az így vizsgálható
kérdések szinte kizárólag az alapvető kognitív mechanizmusok
(memóriaformálás, egyszerű tanulási folyamatok) vizsgálatára
szorítkoznak.
Újabb eredmények nyomán sikerült a családi kutyára
is kiterjeszteni az fMRI-re alapozott neurobiológiai kutatásokat.
Kiderült, hogy a kutya az emberrel való szoros kapcsolata és remek
képezhetősége miatt tökéletes alanya az fMRI-módszertanra alapozott
kísérleteknek. Az elmúlt huszonöt év etológiai kutatásai nyomán
rengeteg ismeretre tettünk szert a kutyák viselkedéséről és kognitív
képességeiről, így ezek neurobiológiai hátterének feltárása új
utakat nyithat a kognitív idegtudományban.
A kutya mint modell
az összehasonlító idegtudományban
Éber kutyák agyi aktivitásának mágneses rezonanciás képalkotási
módszerrel történő vizsgálata az Eötvös Loránd Tudományegyetem
Etológia Tanszék munkatársainak ötlete volt. A 2005-ben kezdődött
kutatás első sikeres felvételei a Kaposvári Egyetem Egészségügyi
Centrumában készültek (Tóth et al., 2009). Már a kezdetekkor fontos
szempont volt, hogy a kutatások fő célja az ember és a kutya
agyműködésének közvetlen összehasonlítása legyen. Ehhez jó alapot
adott az az etológiai megalapozottságú feltételezés, hogy a
háziasítás során a kutyák olyan viselkedési változáson mentek
keresztül, amelyek alkalmassá tették a fajt az emberi közösségekben
való létre. Valójában ma sem tudható pontosan, hogy e változást
milyen genetikai mechanizmusok tették lehetővé, a viselkedés
funkciója terén azonban számos olyan hasonlóság mutatható ki ember
és kutya között, ami alátámasztja e feltételezés jogosságát.
Elképzelésünk szerint a kutya számára a komplex emberi környezet
erős szelekciós hatással bírt, és ez vezetett a viselkedési
konvergenciák kialakulásához. Ennek megfelelően a kutya sok
viselkedési jelleget tekintve jelentősen eltér az őséhez ma is
nagymértékben hasonlító farkastól, hiszen ez utóbbi faj egyedei még
a legerősebb szocializáció esetén sem mutatnak számos specifikus
kutyatulajdonságot.
A jelenleg alkalmazott mágneses rezonanciás
képalkotási eljárások korlátai miatt (lásd alább), a kutya az
egyetlen nagytestű emlős, amely ilyen körülmények között elmélyült
kutatást tesz lehetővé. Ráadásul a kutya őseinek különleges
története külön érdekességet kínál az összehasonlító idegtudomány
számára.
Ma úgy tartjuk, hogy a kutya és az ember közös őse
valamikor 90–100 millió évvel ezelőtt élhetett a Kréta korban
(Springer et al., 2003). A Ragadozók rendjébe sorolt fajok a Kréta
kori tömeges kihalás után jelentek meg, és későbbi evolúciójuk során
is egy jól elkülönült csoportot alkottak különleges életmódjuknak
köszönhetően. Hozzájuk képest elkülönülve jelent meg egy teljesen
független evolúciós vonal, amelynek képviselői a ma élő rágcsálók és
főemlősök egykori elődei voltak. Mindez azt jelenti, hogy a ma élő
kutyák lényegében egy 90–100 millió éves eseménysorozat jelenkori
képviselői, és a különböző evolúciós irányokat figyelembe véve, erre
az időpontra tehető a főemlősök és a ragadozók közös ősének léte is.
Természetesen az eltelt több tízmillió év során az agy mindkét
evolúciós irányban, méretét és szerkezeti felépítését tekintve is
hatalmas változásokon ment keresztül (Lyras, 2009). Ugyanakkor jogos
az a feltételezés, hogy az emberével összevetve mégiscsak a kutya
agya képviseli jobban az ősi formát.
Az összehasonlító anatómia gyakran használ hasonló
megnevezéseket topológiailag megegyező agyterületek elnevezésére. A
kutatók azokban sokat vitatkoznak azon, hogy az ilyen hasonlóság
vajon kellő bizonyíték-e a homológiára, azaz hogy a jelleg egy közös
ősre vezethető vissza. Például mind a kutya, mind az ember esetében
ismert, hogy a temporális lebeny a több millió éves evolúció alatt
lényegesen átalakult, és noha mindkét fajnál leírják a Sylvian
fissure-t (Sylvius-féle oldalsó agybarázda), ez az agyterület a
közös ősben még kevéssé alakult ki. Így pusztán az anatómiai adatok
alapján a két struktúra viszonya nem tisztázható.
Éppen az ilyen problémák felderítésére ad kiváló
lehetőséget az ember és kutya agyműködésének összehasonlítása
képalkotó módszerekkel. Lényeges szempont ugyanis, hogy az agy
vizsgálata során olyan ingereket lehet alkalmazni, amelyeket az
emberi környezetben élő kutya is megtapasztalt, így az újdonság
hatása vagy éppen az attól való félelem szerepe kizárható az egyik
vagy másik faj esetében. Feltételezésünk szerint ha a kutya és az
ember általuk jól ismert ingerekkel találkozik, amelyet hasonló
szabályszerűség szerint dolgoz fel az agyuk egy topográfiailag
megfeleltethető területen, akkor nagy eséllyel homológ struktúráról
lehet szó. Természetesen néhány ilyen felismerés nem elegendő az
általánosításhoz, ugyanakkor az emberi agyra vonatkozó tudásunkra
alapozva a módszer kiváló lehetőséget ad a kutyaagy működésének
feltérképezéséhez, és ezáltal az is érthetővé válhat, hogy egyes
funkciók milyen agyi evolúción mentek keresztül.
Kutyák tréningezése
éber fMRI-vizsgálatokhoz
Az ELTE Etológia Tanszékén közel húsz éve zajlanak kutatások
kutyákon, és büszkék vagyunk arra, hogy ma már nem kell mindent
„megfigyelésnek” hívnunk, mert a hajdan riadalmat keltő „kísérlet”
szó hallatán senki nem gondol olyan módszertanra, amely fájdalmat
vagy komoly stresszt okozhatna a kutyáknak. Nem tartunk
laborkutyákat, hiszen épp akkor vált sikeressé e sokáig
„mesterséges” fajnak tartott különleges háziállat visszacsempészése
az etológia izgalmas alanyainak sorába, amikor a tudományos
közvélekedés is elfogadta, hogy a kutya természetes szociális
környezete az emberi csoport.
Az interspecifikus kommunikációtól a szociális
tanuláson át az agresszióig, kivételesen széles palettán folytatunk
vizsgálatokat, de mindig figyelünk arra, hogy ne ártsunk a
kutyáknak, azaz olyan módszerekkel teszteljük a viselkedésüket,
amelyek az ő jólétüket is figyelembe veszik. Ilyen megközelítésből
különösen komoly kihívást jelentett közel tíz évvel ezelőtt az az
ötlet, hogy a viselkedés elemzése mellett pillantsunk bele a kutyák
neurális működésébe is. A laboratóriumi állatok agyi feldolgozó
folyamatainak vizsgálatára kidolgozott klasszikus módszerek szinte
kizárólag invazív módszereket használnak, ami addig járatlan utak
felfedezésére inspirált minket.
Míg a modern képalkotó eljárások kifejlesztése az
ember esetében új távlatokat nyitott, nem véletlen, hogy a
szocio-kognitív képességek reprezentációjában szerepet játszó agyi
mechanizmusok fMRI-vel való feltérképezésére állatok esetében alig
nyílt lehetőség. Számos nehézséggel kell számolni, amelyek
megnehezítik az agyi aktivitás feltárását; az MRI-készülék által
keltett hangos, folyamatos zaj, illetve a rezgés már önmagában is
kellemetlen, ráadásul a funkcionális mérések módszertana
megköveteli, hogy a vizsgálati alany feje a felvétel alatt
tökéletesen mozdulatlan legyen. Állatok esetében nagyobb a
valószínűsége a mozgási műtermékek keletkezésének, amelyek a mérési
adatok kiértékelését nehezítik, sokszor lehetetlenné teszik. Ez
emberek vizsgálatakor is jelent néha gondot, különösen mert egy idő
után kissé kényelmetlenné válhat az adott testhelyzet, és ez
elmozdulásra készteti az alanyt. Állatoknál további problémát
okozhat, ha az alany a teszt során alkalmazott ingerre mozgással
(például ingerforrás keresése, attól való elhúzódás) igyekszik
válaszolni. E metodikai nehézségeket az eddigi vizsgálatok
legtöbbjében az állat kábításával vagy rögzítésével oldották meg. A
kábítás fő hátránya, hogy a beadott szer hatására kimutathatóan
csökken az agykérgi aktiváció mértéke (Lahti et al., 1999), így
ugyanarra az ingerre gyengébb vagy módosult választ kaphatunk. A
rögzítés pedig amellett, hogy a helyzet kényszerítő jellege még
intenzív habituáció ellenére is befolyásolhatja az agyi választ
(Zhang et al., 2000), számunkra állatvédelmi/etikai szempontból is
elfogadhatatlan (az állat fejét a készülékhez rögzítik, így ha
elmozdul a kívánt testhelyzetből, azonnal fájdalmat érez, tehát
nincs, vagy csak korlátozott mértékben van lehetősége egy adott
ingerre eltávolodással reagálni).
Egyértelmű volt tehát, hogy amennyiben éber és
rögzítetlen kutyákat szeretnénk vizsgálni, csak egy teljesen új
módszer kialakításával érhetünk célt, ezért speciális tréningtervet
dolgoztunk ki. Mivel minden kutatásunknál lényeges szempont, hogy
lehetőleg reprezentatív mintát vizsgáljunk (így könnyebben
általánosíthatók az eredményeink), olyan módszer alkalmazására
törekedtünk, amely átlagos családi kutyák esetében is beválik, nem
csupán egy kivételesen képzett vagy speciális temperamentumú, szűk
csoportnál.
A tréning végső célja az volt, hogy a kutyák
hét-nyolc percig éber állapotban, hason fekve, fejüket két mellső
lábuk közt az asztalra fektetve, mozdulatlanul feküdjenek az
MRI-készülékben. A mozdulatlanul szót ez esetben egészen szigorú
értelemben kell venni, mert ha az orrukat megnyalják, vagy egy
nagyobbat nyelnek már az elfogadható mértéknél több lesz a mozgás a
felvételen. Ebben az esetben a mérés sikertelen, hiábavaló
próbálkozásnak minősül.
A tréningprogram a gazdák bevonásával és gyakorlott
kiképzők irányításával zajlott egy lépcsőzetesen egymásra épülő
fázisokat tartalmazó protokoll szerint. Az első szakaszban a kutyák
egy külső helyszínen megtanultak asztalon „hasalni”, hozzászoktak
ahhoz, hogy az asztal mozog alattuk, és hogy magnóról az
MRI-készülék hangját játsszuk vissza nekik. A készülék zajának
tompításához és a hangingerek adásához használt fülhallgató,
valamint a tekercset tartó pánt viselését is külön tréningeztük. A
képzés ezen első szakaszában pozitív megerősítésen alapuló
technikákat alkalmaztunk (például klikker-tréninget [Pryor, 2002]).
Ha a kutyák
|
|
|
már percekig megbízhatóan hasaltak, megkezdődhetett
az MRI-gép melletti tréning, amelynek célja, hogy teljesen nyugodtan
viselkedjenek a későbbi tesztek helyszínén. A legtöbb etológiai
kísérlettel szemben, ahol a kutyáknak labdák és finom falatok
megszerzésére nyílik lehetőségük, az fMRI-tesztek nemcsak zajosak,
de kissé unalmasak is, amit nem feltétlenül ellensúlyoz némi
jutalomfalat a teszt legvégén. Ezért ebben a szakaszban az
elsődleges célunk az volt, hogy a kutya ennek ellenére élvezze a
részvételt, fontos legyen neki, hogy a gazda kérésére ott hasal az
MRI-készülékben, és figyeli az ingereket. Ezt szociális tanuláson
alapuló módszerek alkalmazásával értük el. A kiképzési módszer
lényegi eleme az ún. rivális tréning (McKinley – Young, 2003). Ez
azon a jelenségen alapul, hogy a szociális fajok egyedei spontán
(közvetlen jutalom nélkül) tanulnak társaik viselkedését
megfigyelve. Jelen esetben igyekeztünk egy másik, ismerős kutyát
tréningezni, és közben látványosan dicsérni, jutalmazni a
MRI-készülékben, mialatt a kezdő kutya szabadon járkálhatott a
szobában, és senki sem foglalkozott vele. A kezdő kutyák ebben a
helyzetben jellemzően igyekeztek abba a pozícióba kerülni, ahol a
társukat látták, így alig várták, hogy rájuk kerüljön a sor. Ha a
kutya lelkesen elfoglalta a helyét a MRI-készülékben, és maga
ajánlotta fel a hasalás pozíciót, már könnyebb volt a 7–8 perces
mozdulatlanságot elérni. A tréningek és a tesztek alatt is
elsődleges szempont volt, hogy a kutyák bármikor azonnal ki tudták
húzni a fejüket a pánt alól, és kijöhettek a MRI-készülékből, azaz
minden kutya csak a gazda kérésére maradt fekve, erre semmi sem
kényszerítette.
Korai fMRI-vizsgálatok kutyán
Az elmúlt években egy egyesült államokbeli kutatócsoport, Greg Berns
és munkatársai (2012) is sikeresen alkalmaztak fMRI-t kutyák
vizsgálatára. Azt találták, hogy a jutalmat előrejelző kézjelzés egy
semleges kézjelzéssel szemben aktivitáserősödéshez vezet az agyi
jutalomközpontként számon tartott nucleus caudatusban. Ez a
vizsgálatsorozat azonban több szempontból is kritizálható. Egyrészt
összesen két kutyával dolgoztak, tehát nem volt lehetőség
csoportszintű elemzésre, így valódi populációs szintű
következtetésekre sem. Másrészt az általuk vizsgált kutyák nem
feküdtek végig mozdulatlanul a mérés során. A kutatók a mozdulatlan
periódusok kiemelésével, és csak ezek elemzésével igyekeztek
kiszűrni a mozgásból származó zajt, de könnyen lehet, hogy ez is
hozzájárult ahhoz, hogy a kapott eredményeik nem voltak robosztusak.
Harmadrészt, Berns és munkatársai igyekeztek párhuzamokat vonni az
emberi és a kutyaagy működése között, de ezt anélkül tették, hogy
vizsgálatukat emberi alanyokon is elvégezték volna. Egyetértünk
azzal, hogy az ember–kutya összehasonlítás, az evolúciós kontextusba
helyezés lehetősége teheti a kutya fMRI-módszerét igazán relevánssá,
de azt gondoljuk, hogy az összehasonlító vizsgálatok egyik
alapfeltétele, hogy ugyanazt a kísérletet, lehetőleg azonos
ingerekkel és körülmények között végezzük el a két fajon. A családi
kutyák számára nagyrészt ugyanazok az ingerek relevánsak és
ismerősek, mint az ember számára, ami megkönnyíti az
összevethetőséget. Az itt bemutatott tréningezési módszerünkkel
pedig az is lehetővé vált, hogy a kutyák hosszú perceken át
mozdulatlanul feküdjenek, ezért a kísérleti elrendezés is közel
azonos lehet a két faj vizsgálatakor.
Az első kutya–ember összehasonlító
fMRI-vizsgálat és eredményei
Az fMRI-vizsgálatunk fő célja az volt, hogy funkcionális analógiákat
keressünk az ember és a kutya hallókérge, hangfeldolgozó neurális
mechanizmusai között (Andics et al., 2014). A hallórendszer fontos
szociális funkciója a hangadó azonosítása és érzelmi állapotának
feldolgozása, vagyis, hogy megállapítsa az egyed kilétét és
hogylétét. Fajtárshangokra való kitüntetett érzékenységet találtak
magasabb szintű hallókérgi régiókban az emberi agyban (Belin et al.,
2000) és más főemlősök agyában is (Petkov et al., 2008). Korábbi
humán kutatások kimutatták egyes hallókérgi régiók szerepét a
hangokban rejlő érzelmi információk feldolgozásában is. Nem
főemlősök hasonló agyi funkcióiról azonban eddig nagyon keveset
tudtunk. Pedig a fajtársak és nem fajtársak hatékony elkülönítése,
hasonlóan az észlelt érzelmi állapotok megkülönböztetéséhez, a
főemlős és nem főemlős fajok számára egyaránt alapvető fontosságú
olyan döntési helyzetekben, mint a párválasztás, a territórium
védelme vagy a hierarchiához kapcsolódó kihívások.
Pontosan ugyanazt a neurális képalkotási kísérletet
végeztük el ember és kutya résztvevőkkel – ilyen vizsgálatra
korábban még nem volt példa. Az emberek és a kutyák is éberen,
lekötözés nélkül vettek részt a kísérletben, mozdulatlanul feküdtek
a vizsgálóasztalon három hatperces mérés során. Megmértük a kutyák
és az emberek agyi aktivitását, miközben azok összesen közel kétszáz
kutya- és emberi hangot hallgattak meg. A hangok érzelmi töltete
parametrikusan változott, volt köztük nagyon negatív és nagyon
pozitív is, a repertoár a nyüszítéstől és zokogástól egészen a
játékos csaholásig és nevetésig terjedt. Egyéb ismerős, de
nemvokális környezeti zajokat (például harangzúgás, motorhang) is
bemutattunk az alanyoknak.
A hangokra érzékeny területeket funkcionálisan
azonosítottuk mindkét fajban (a bármilyen hang vs. csend kontraszt
segítségével). Hasonló kérgi és kéreg alatti területeket
azonosítottunk (perisylvian régiók, mediális genikulátusz) kutyánál
és embernél, viszont a területek összesített térfogata lényegesen
különbözött a két fajnál: kutyában 12 cm3, emberben 95 cm3
hangokra érzékeny területre szűkítettük le a további vizsgálódás
keresési terét.
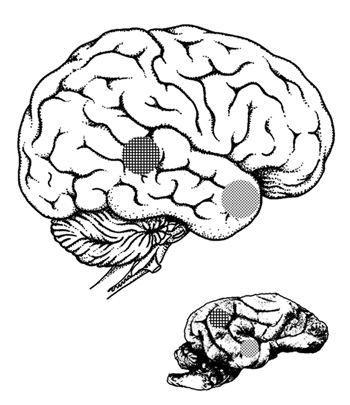
1. ábra • Hasonló funkciójú hallókérgi
területek
az ember és a kutya agyában. Pontozott régió
(temporális pólus): fajtárshangokra való érzékenység;
Négyzetrácsozott régió (korai hallókérgi terület): az emberi és
kutyahangok érzelmi töltetére való érzékenység.
(A méretkülönbségek körülbelüli arányt jeleznek.)
A mérések szerint a fajtárshangokra érzékeny
agyterület (ún. voice area) kutyában is és emberben is közel
ugyanott, a halántéklebeny elülső végében, az ún. temporális
pólusnál helyezkedik el. Ez az eredmény nagy valószínűséggel azt is
jelzi, hogy a fajtárshangokra való érzékenység egy olyan agyi
funkció, ami már legalább százmillió éve, a kutya és az ember
legutolsó közös őse óta jellemzi az emlősök hangfeldolgozását. Ez a
terület a kutyában elsősorban a kutyahangokra, emberben pedig az
emberhangokra reagál, de nem mutat erősebb érzékenységet a másik faj
vokalizációira, a környezeti zajokhoz képest. Emberben a
halántéklebeny hátulsóbb területei (mediális és posterior superior
temporalis sulcus) is az emberi hangokra reagáltak legerősebben, de
az elülső területekkel szemben a kutyahangokat is preferálták a
környezeti zajokkal szemben. Ez azt mutatja, hogy a humán hátulsó
területek nemcsak a fajtársak hangjaira érzékenyek, hanem általában
az ismerős vokalizációkra is.
Megfigyeltünk egy nagyon izgalmas hasonlóságot
abban is, ahogy a kutya és az ember agya a hangokban rejlő
érzelmeket feldolgozza. A legmeglepőbb, hogy a kutya agyában is
ugyanaz a korai hallókérgi terület reagál erősebben a pozitívabb
hangokra, mint az emberében – ráadásul ez független attól, hogy
melyik faj hangjairól van szó. Korábbi kutatásaink fényében (Faragó
et al., 2014) ez az eredmény arra is utalhat, hogy mindkét faj
ugyanazokat az egyszerű akusztikus információkat (hanghossz,
alapfrekvencia) használja a hangokban rejlő érzelmek kódolására.
Ennek az akusztikai feldolgozási folyamatnak egy állomását jelezheti
ez a korai, fajok közt is nagyon hasonló hallókérgi érzékenység. A
jelen eredmény lehet az első közvetlen bizonyíték arra, hogy az
emlősök hangfeldolgozási mintázata jól követi a vokalizációk
strukturális-funkcionális szerveződését, ez pedig megalapozza
bizonyos akusztikus paraméterek felhasználását a fajok közti
hangfelismerés folyamatában.
Ez a kutatás az első lépés ahhoz, hogy megértsük,
milyen agyi mechanizmusok révén képes a kutya hatékonyan
ráhangolódni a gazdája lelkiállapotára, és mitől tájékozódik olyan
jól a mi szociális környezetünkben. Szélesebb értelemben kutatásunk
segít feltárni azokat a viselkedési és neurális mechanizmusokat,
amelyek a két faj több tízezer éve fennálló szövetségét olyan
hatékonnyá tették. Ezek az első eredmények azt mutatják, hogy
nemcsak az ember és a családi kutya szociális környezete hasonlít,
hanem azok az agyi mechanizmusok is, amelyek segítségével a
szociális információ feldolgozásra kerül. Az ún. voice area megléte
mindkét fajban és az érzelmekre mutatott hasonló topográfiájú agyi
aktivitás jól magyarázza a két faj közti kommunikáció sikerességét.
Ugyanakkor érdekes módon, míg az ember hallókérgi
területeinek csak három százaléka aktiválódik erősebben a nem
élőlénytől származó zajokra, kutyáknál ez az arány 48 százalék.
Vagyis míg az emberi hallókéreg szinte egészében az emberhangokra
optimalizálódott, a kutya hallókérge ennél sokkal heterogénebb
működésű. A fajtárshangokra érzékeny területek az emberben fontos
szerepet játszanak a nyelvi ingerek feldolgozásában is. Érdekes
kérdés, hogy az emberi beszédértést segítő agyi mechanizmusok közül
melyek vannak jelen a kutyában is, és ezek milyen beszédértési,
nyelvi képességekkel ruházzák fel az állatot. További kutatásaink
egyik fő csapásiránya ennek a kérdésnek a megválaszolása lesz.
Kulcsszavak: mágneses rezonanciás képalkotás, összehasonlító
idegtudomány, fajtárshangok, érzelemészlelés, fMRI
IRODALOM
Andics Attila – Gácsi M. – Faragó T. et
al. (2014): Voice-sensitive Regions in the Dog and Human Brain Are
Revealed by Comparative FMRI. Current Biology. 24, 5, 574–578. DOI:
10.1016/j.cub.2014.01.058
Belin, Pascal – Zatorre, R. J. – Lafaille,
P. et al. (2000): Voice-selective Areas in Human Auditory Cortex.
Nature. 403, 6767, 309–312. DOI: 10.1038/35002078 •
WEBCÍM
Berns, Greg S. – Brooks, A. M. – Spivak,
M. (2012): Functional MRI in Awake Unrestrained Dogs. PLoS One. 7,
e38027. DOI: 10.1371/journal.pone.0038027 •
WEBCÍM
Faragó Tamás – Andics A. – Devecseri V. et
al. (2014): Humans Rely on the Same Rules to Assess Emotional
Valence and Intensity in Conspecific and Dog Vocalizations. Biology
Letters. 10, 1, 20130926. DOI: 10.1098/rsbl.2013.0926 •
WEBCÍM
Lahti, Katariina M. – Ferris, C. F. – Li,
F. et al. (1999): Comparison of Evoked Cortical Activity in
Conscious and Propofol-anesthetized Rats Using Functional MRI.
Magnetic Resonance in Medicine. 41, 2, 412–416. DOI:
10.1002/(SICI)1522-2594(199902) 41:2<412::AID-MRM28>3.0.CO;2-3 •
WEBCÍM
Logothetis, Nikos K. (2008): What We Can
Do and What We Cannot Do with fMRI. Nature. 453, 7197, 869–878.
DOI:10.1038/nature06976 •
WEBCÍM
Lyras, George A. (2009): The Evolution of
the Brain in Canidae (Mammalia: Carnivora). Scripta Geologica. 139,
1–93. •
WEBCÍM
McKinley, Sue – Young, Robert J. (2003):
The Efficacy of the Model–rival Method When Compared with Operant
Conditioning for Training Domestic Dogs to Perform A
Retrieval–selection Task. Applied Animal Behaviour Science. 81, 4,
357–365. DOI: 10. 1016/S0168-1591(02)00277-0 •
WEBCÍM
Petkov, Christopher I. – Kayser, C. –
Steudel, T. et al. (2008): A Voice Region in the Monkey Brain.
Nature Neuroscience. 11, 3, 367–374. DOI:10.1038/nn2043
Pryor, Karen (2002): Clicker Training for Dogs. Ringpress Books Ltd,
Tóth Lilla – Gácsi M. – Miklósi Á. et al. (2009): Awake Dog Brain
Magnetic Resonance Imaging. Journal of Veterinary Behavior: Clinical
Applications and Research. 4, 2, 50. DOI: 10.1016/j.jveb.2008.09.021
Van der Linden, Annemie – Van Camp, N. –
Ramos-Cabrer, P. et al. (2007): Current Status of Functional MRI on
Small Animals: Application to Physiology, Pathophysiology, and
Cognition. NMR in Biomedicine. 20, 5, 522–45. DOI: 10.1002/nbm.1131
•
WEBCÍM
Zhang, Zhiming – Andersen, A. H. – Avison,
M. J. et al. (2000): Functional MRI of Apomorphine Activation of the
Basal Ganglia in Awake Rhesus Monkeys. Brain Research. 852, 2,
290–296. DOI: 10.1016/S0006-8993(99)02243-X
|
|
