Növényökofiziológia
Az autökofiziológia
és szünfiziológia diszciplináris
helyzete az ökológián belül
Tuba Zoltán
DSc, mb. intézetigazgató, egyetemi tanár
SZIE Növénytani és Ökofiziológiai Intézet, MTA–SZIE Növényökológiai Kutatócsoport
tuba . zoltan ![]() mkk . szie . hu
mkk . szie . hu
A növényökofiziológia ökológiai diszciplína. Az ökológián belül pedig a működést leíró funkcionális ökológia része, amely a növényközösség/ek vagy ökoszisztémáik működésének fiziológiai folyamatait tanulmányozza.
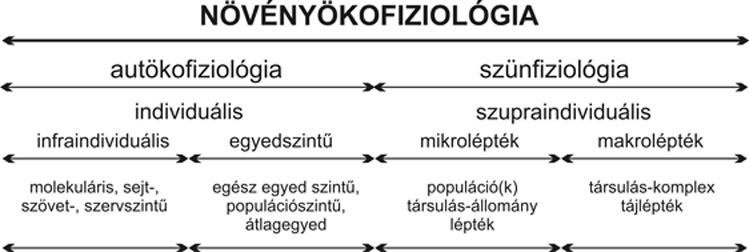
Szerveződési szint szerint a növényökofiziológián belül a) egyed- vagy autökofiziológiát és b) közösségi vagy szünfiziológiát kell megkülönböztetnünk. (A szün, eredeti írásmódban syn szócska, görög elöljáróból alakult előtag, jelentése együtt, egyszerre, ami itt a közösségi, egyedfeletti vagy szupraindividuális jellegre utal.) Amíg az autökofiziológia a növényközösség populációra reprezentatív egyedeinek fiziológiai válaszait vizsgálja, addig a szünfiziológia a növényegyedekből szerveződő közösség (Fekete, 1998) kollektív, szupraindividuális fiziológiai válaszait kutatja. Fogalmazhatunk továbbá úgy is, hogy az autökofiziológia az infraindividuális és individuális léptékű, a szünfiziológia pedig a szupraindividuális léptékű részdiszciplína.
A szünfiziológia célja a növényközösségek – a mindenkori adott kényszerfeltételek (limitációk) közötti – állomány-, ökoszisztéma és tájléptékű fiziológiai folyamatainak és azok szabályozásának a leírása (Tuba et al., 1998a). A szünfiziológia tehát nem teljesen fedődik a növényökofiziológia egészével, hisz annak egy (jó) része autökológiai, individuális és infraindividuális jellegű és megközelítésű. Éppen előzőek miatt lényeges és szükséges a szünfiziológia elkülönített, az ökofiziológia önálló részdiszciplinájaként való bevezetése.
Nem sokszor hangzott el eddig még, hogy a populáció egyaránt tanulmányozható infra- és szupraindividuális szinten, vagyis autökológiai és szünfiziológiai szemlélettel is: ugyanis az átlagegyeddel reprezentált populáció az autökofiziológia, míg az egyedek közösségeként, állományaként kezelt és vizsgált populáció pedig a szünfiziológia területére esik.
Szünfiziológiai minden olyan működési folyamat, ami a növénytársulás és ökoszisztémája – mint egyed és az átlagegyeddel reprezentált populáció feletti szupraindividuális szerveződésű rendszer – egészének anyag- és energiafelvételét, átalakítását, tárolását és leadását, azaz egy termodinamikai nyílt rendszer és környezete közötti anyag- és energiaáramlást végzi és szabályozza. Különös jelentőséget ad a növényközösségek (társulások) (szün)fiziológiájának, hogy az az ökoszisztémák egészének a létét is meghatározza.
Mivel a növénytársulások létezésének (energiafelvételének, növekedésének, fejlődésének, szervesanyag-termelésének stb.) (is) a fotoszintetikus CO2-asszimiláció és a C-metabolizmus az alapja, ezért a szünfiziológia a társulás-fotoszintézist és a társulás C-metabolizmust és mindezek limitációját kezeli kitüntetetten. Mindez pedig tükröződik a társulásfiziológia módszertárában is. A társulások fiziológiai működése mindig egy vagy inkább több tényező korlátozó kényszere mellett megy végbe, ezért a szünfiziológia a társulás fiziológiai folyamatait a társulásban éppen limitáló tényezőkkel (vízhiány, alacsony vagy magas hőmérséklet, tápanyaghiány, kompetíció, rovargradáció, tűz stb.) összefüggésben vizsgálja. A fiziológiai működés a reproduktív allokáción keresztül szorosan kapcsolódik a társulás működésének másik fontos oldalához, a fajok populációinak szaporodásdinamikájához, és így a populációdinamikához, melyen keresztül közvetve szerepet játszik a társulások fajösszetételének, faj/egyed diverzitásának, szerkezetének és produkciójának a kialakításában és a vegetációdinamikai folyamatokban.
A társulásfiziológiai (ún. szünfiziológiai) folyamatok/sajátosságok létezését és a szünfiziológia művelésének szükségességét már önmagában az is indokolja, hogy a növénytársulások és ökoszisztémáik szintjén a fiziológiai folyamatok sajátosságai mások, mint annak növényegyedeiben és különösen azok szerveiben, szöveteiben és sejtjeiben, illetve hogy vannak csak a társulás egészére jellemző, az egyedek és infraindividuum szintjén nem megjelenő fiziológiai sajátosságok is. Így például a társulás zöld lombsátrának egésze – csupán a levélszögállások, levélorientációk és a vertikális rétegzettség miatt is – teljesen más tulajdonságú, optikai szerkezetű fotoszintetikus apparátus, mint a társulás egyes egyedeinek hajtásai vagy még inkább az egyes leveleké és sejteké. Ezért a növénytársulásban a szintezettség kialakulásával fokozódik a záródás, nő a társulás egységnyi területére jutó összlevélfelület (LAI), és csökken a megvilágítás, amely csökkenti ugyan az egyes egyedek egységnyi levélterületére eső fotoszintézis-intenzitását, azonban a LAI említett növekedése, valamint a lombozatarchitektúra és az extinkciós koefficiens egyidejű változása miatt az egységnyi társulásterületre eső összfotoszintézis teljesítménye mégis nő.
A szünfiziológiai folyamatok időléptéke a fiziológiai folyamatokétól nagyobb, illetve az ennél kisebb időléptékű folyamatok nagyobb időintervallumokban integrálva nyernek szünfiziológiai és ökológiai értelmet. Hovatovább már az egyedek ökofiziológiájánál is csak a nagyobb időtartam alatt lezajlott folyamatok integrálása révén lehet az egyed viselkedését magyarázni. Az egész egyed szempontjából pedig leginkább az az időtartam (minimálisan 24 óra) tekinthető ökológiailag relevánsnak, amely során például a fiziológiai folyamatok már mérhető változást eredményeznek az egyedek szervallokációs mintázatában.
A növényökofiziológia és két ága, az autökofiziológia és szünfiziológia diszciplináris helyzetének tisztázásával sikerült feloldanunk azt a megértést sokszor zavaró fogalmi sokféleséget (növényökofiziológia, közösségi élettan [community physiology], szisztém- vagy rendszerélettan [system physiology], környezeti növényélettan, ökológiai növényélettan, élettani ökológia, stresszélettan stb.), ami a növényökofiziológiát körüllengte. Előzőek közül külön is meg kell említenünk a növényi stresszélettant, amely csak és kizárólag akkor tekinthető növényökofiziológiai (főként autökofiziológiai) szempontból relevánsnak, ha az a stressztoleranciát (is) vizsgálja, és hogyha az a szupraindividuális (közösségi, társulási, ökoszisztéma) szerveződésű és léptékű folyamatok megértéséhez is egyértelműen hozzájárul. Ugyanis a növényi stresszélettant igen gyakran a növényökofiziológia szinonimájaként avagy egyértelműen a növényökofiziológia részeként említik, pedig fentiek alapján nyilvánvaló, hogy nem ez a helyzet. Annak ellenére nem, hogy a növényi stresszfiziológia a környezeti tényezők limitációinak a vizsgálatával történetileg először az ökológiában, éppen az ökofiziológia részeként jelent meg (Lösch – Larcher, 2000). A stressz egyébiránt a természetes élőhelyeken a növényközösségek és ökoszisztémáik egészét érintve szupraindividuálisan is megjelenik.
Autökofiziológia versus szünfiziológia: antitézis vagy komplementer szükségesség, avagy szükség van-e az autökofiziológiára?
A növényökofiziológia két ága valójában ugyanazon ökológiai kérdések két eltérő szerveződési szint szerinti megközelítésének leképződése: az autökofiziológia az organizmus, az egyedi szervezet élettani és morfológiai adaptációja felől közelítve elemzi az ökológiai problémát, a szünfiziológia pedig a közösség, a szupraindividuum, a társulás és az ökoszisztéma egésze felől közelít.
A környezethez történő alkalmazkodás – összhangban a multiplurális környezetelvvel (Juhász-Nagy, 1986) – a molekuláristól a sejt-, szövet-, szervszinten át az egyedig és populációig sokféle szinten bekövetkezhet. Vagyis minden olyan szinten történhet adaptáció, ahol a környezeti tényezők detektálása zajlik és bekövetkezik a válasz a környezeti tényezők változására. Az adaptációban az élettani és molekuláris folyamatokon kívül főként azoknak van komoly szerepük, amelyek genetikailag meghatározottak, de kifejeződésük erősen függ a környezeti tényezőktől. Mivel a növények helyhez kötöttek, számukra a fiziológiai és morfológiai adaptáció különösen fontos. A környezeti tényezők növénnyel való kölcsönhatásai alakítják ki az adott növény „fitneszét”, azaz a növekedés és szaporodás képességének a sikerét egy adott időben és térben.
Mindezek tanulmányozása pedig az autökofiziológia tárgykörébe tartozik (Larcher, 2003). Az autökofiziológia tehát nélkülözhetetlen a szupraindividuális fiziológiai működés megértéséhez. Így például növényegyedek reproduktív stratégiája, reproduktív allokációja egyaránt vizsgálható autökológiai és szünfiziológiai szinten is. Addig, amíg az autökológia mint adaptációs formát elkülönítve vizsgálja a fenti példánál maradva a reproduktív allokációt, addig a szünfiziológia rendszerbe (egyedek rendszerébe) ágyazva vizsgálja ugyanezt az allokációs problémát. Előzőeknek messzemenő következményeik vannak a közösségek egészének fiziológiai működésére, nélkülük a szünfiziológiai folyamatok nem érthetők meg teljesen. Épp ezen előzőek miatt mondhatjuk azt, hogy a növényökofiziológia jellegzetesen olyan ökológiai diszciplinaág, amely a nagy tér- és időléptékű módszerek mellett a kezdetektől alkalmaz biokémiai, biofizikai, sőt molekuláris módszereket is, éppen a szupraindividuális működés megértése érdekében. Hovatovább az ökofiziológia nemcsak molekuláris módszereket használ, hanem az ökofiziológia, az autökofiziológia a molekuláris biológia fejlődését is előmozdíthatja, mivel jól felhasználható bizonyos élettani és morfológiai tulajdonságok alkalmazkodásának vizsgálatában, ami még ráadásul populációgenetikai és az evolúcióbiológiai vonatkozásokkal is bír.
Noha úgy tűnik, a növényökofiziológia szemléletileg ma egyre inkább a szupraindividuális léptékű szünfiziológia irányába halad, mégis módszertanilag az autökofiziológia is mindig megújul, (például stabil izotópok [C, O, N], fluoreszcenia leképezési, stabil izotópok frakcionálódása és diszkriminációja, avagy éppen a molekuláris genetikai módszerek használata révén). Ez persze azért is fontos, mert nem kis számban az autökofiziológia módszertárából fejlődnek ki a szünfiziológiai vizsgálatokra alkalmas metodikák (például szén-dioxid- és vízgőzcsere mérése a növényállomány szintjén) is.
A szünfiziológiának nagy szüksége van az egész egyedek (az egyes egész egyedek) fiziológiájának ismeretére is. Ugyanis a szünfiziológiai válaszok és folyamatok kumulatív jellegűek. A mérésükre szolgáló módszerek/technikák nem tájékoztatnak az alkotó egyedek fiziológiai válaszairól.
Az egész egyedszintű fiziológiai folyamatok területén a továbblépéshez viszont az egész egyedek mérésére alkalmas technikák kifejlesztése szükségeltetik. Ugyanis jelenleg az autökofizológiai vizsgálatokhoz szükséges műszerek szinte mindegyike laboratóriumi telepített műszerek hordozhatóvá és így terepi körülmények között használhatóvá tett változatai, melyek éppen „infraindividuális” eredetük miatt jó esetben is legfeljebb egész levél mérését teszik lehetővé, és sajnos még egy kisméretű lágyszárú növény egész egyede sem mérhető meg.
Mindezek következménye, hogy napjainkban a növényegyedek egészének fiziológiájára vonatkozó ismereteink valójában még igen szerények. Ezért amikor egész növényegyed fiziológiájáról (egyed autökofiziológiájáról) beszélünk, azon ma döntően egyed alatti, infraindividuális (levél/hajtás, gyökér, reproduktív szerv stb.) szintű mérésekből származó fiziológiát kell értenünk. Vagyis az egész növényegyedekre vonatkozó fiziológiai ismereteink ma többnyire csupán az egyes szervek válaszainak az egész egyedre vetített kiterjesztéséből (egyszerű felszorzásából) származnak. Az „egész növényegyed” ökofiziológiai folyamatainak megértését persze sokszor az is hátráltatja, hogy nem világos, mit tekinthetünk egyáltalán egyetlen növényegyednek, például a zsombékképző, illetve a klonális növekedésű növények esetében.
Tehát megállapítható, hogy noha a szünfizológia a szerveződési szintek közül a szupraindividuális (társulásállomány) szintet tünteti ki, annak megismeréséhez és megértéséhez az egyedek fiziológiájának ismeretét sem nélkülözheti. Már csak azért sem, mert a szünfiziológiai jelenségek és mechanizmusok megértéséhez vissza kell nyúlnunk az alatta lévő (autökofiziológiai) szerveződési szinthez.
A szünfiziológia mikro- és makroléptéke
A szünfiziológián belül a megismerés eddig a makroléptékű, vagyis az társulásállománytól a tájlépték szintjéig terjedő – fiziológia területén haladt előre (lásd később). Az autökofiziológiai és a fenti makrolépték szünfiziológiája közötti – néhány m2-es ún. mikro-léptéktartományba eső szünfiziológiai működésről viszont gyakorlatilag a legutóbbi évig nem voltak ismereteink. Pedig ez az a léptéktartomány, ahol a növényközösségekben a legtöbb és leglényegesebb folyamat megy végbe (Bartha, 2004). A szünbiológia jellegéből következően a szünfenetikai és szünfunkcionális, így utóbbin belül a szünfiziológiai folyamatokkal szoros kapcsolatban kell álljanak. Ennek ellenére a szünfenetikai és a szünfiziológiai folyamatok tényleges összekapcsolása, azonos léptékű, párhuzamos vizsgálata mégis szinte máig váratott magára. Ennek oka pedig egyszerűen az említett léptéktartományban végzendő szünfiziológiai vizsgálatok hiánya, ami elsősorban az ezen folyamatok mérésére alkalmas módszerek hiányára vezethető vissza. Kutatásainkkal – többek között – a mikroléptékű növényi szünfiziológia fent hiányolt elméleti kereteit és metodikáját alapoztuk meg. Az elméleti alapozás eredményeként a növényökofiziológia alábbi felosztására teszünk javaslatot (Tuba, közöletlen anyag; 1. táblázat):
Az autökofiziológia és makrolépték közötti fiziológia: a mikroléptékű szünfiziológia
Csak a mikrotérléptékű szünfiziológiai vizsgálatokkal adhatók meg a válaszok olyan alapvető fontosságú kérdésekre, mint például:
• létezik-e és ha igen, mekkora a szünfiziológiai minimumarea az egyes növénytársulásoknál?
• van-e összefüggés a cönológiai és a szünfiziológiai minimumarea között?
• befolyásolják-e a domináns és ritka növényfajok, egyáltalán a florisztikai kompozíció a társulásfiziológiai folyamatokat?
• a térben és időben változó cönológiai viszonyok szupraindividuális szinten hogyan befolyásolják a társulásfiziológiai folyamatokat?
A szünfiziológia és ezen belül főként a mikroléptékű szünfiziológia sajátossága, hogy a felsorolt és fel nem sorolt alapkérdésekre a válaszokhoz az ugyanazon léptékű szerkezeti, cönológiai, illetve fajkompozíció, vagyis szimultán szerkezeti ökológiai vizsgálatok is szükségeltetnek (Mooney et al., 1987). Mindezért a szünfiziológiai szemlélet nemcsak újfajta gondolkodást, de új módszereket is kíván. Így a szünfiziológia fejlődésének is egyik záloga a megfelelő, térben és időben adekvát léptékben dolgozó szünfiziológiai módszerek kifejlesztése. Erre minden olyan módszer/technika megfelelő, amely az egész növényközösség, mint sok faj számos egyedéből álló rendszer energia-, hő-, víz- és egyéb anyagfelvételének, átáramlásának, tárolásának, leadásának, metabolizmusának, növekedésének, fejlődésének, produkciójának stb. in situ, többnyire nem destruktív módon történő mérésére alkalmas.
A makroléptékű, több kilométeres vagy ennél is sokkal nagyobb, már tájléptékű fiziológiai működés vizsgálatára használható technikák eredetileg más tudományágak, így a légkörfizika, repülésirányítás és meteorológia részére lettek kifejlesztve. Bármily furcsa is, de előnyük egyben a hátrányuk is, vagyis hogy csak a legnagyobb térléptékű folyamatok vizsgálatára alkalmasak. Ilyen módszer a vízgőz, hő, CO2 és egyéb, pl. további üvegházhatású gázok (nitrogén-oxidok, metán) fluxusainak „eddy korrelációval” hosszú időtartamon keresztüli mérése a társulás és a légtér határán (Nagy et al., 2007). Az ismert metodikák tárházát napjainkban egyre inkább kiegészítik a szünfiziológiailag is releváns távérzékeléses módszerek (például LAI, NDVI, GIS) és természetesen a modellezés (szimulációs, optimalizációs), amely az előrejelzés lehetőségével már a gyakorlati (például környezetvédelmi, tájökológiai) alkalmazást is előkészíti.
Amíg az autökofiziológia-léptékű és a makrotérléptékű módszertannál és ismereteknél nagy az előrelépés, a cönológiailag releváns néhány négyzetméteres mikrotérléptékű szünfiziológia vizsgálata módszertanilag napjainkig megoldatlan volt. Elvileg az ezen léptéktartományú társulásfiziológiai folyamatok mérésére is lett volna lehetőség, ez pedig például az ún. „megnövelt kamrás” CO2 gáz- és H2O gőzcseremérési módszer. Ez a módszer a levél mérésére használt kamrák felnagyított változata. Azonban a levélmérésre alkalmas kamrák méretének egyszerű megnövelése (felnagyítása) nem járható út. Ugyanis a nagy űrtartalmú mérőkamrában számtalan (például légáramlástani, hőmérsékletváltozási és hőmérsékletszabályozási, vízgőzkondenzációs, rövid időtartam alatt használható mérési jel és jelátalakítás stb.) problémát kell megoldani, ráadásul olyanokat, amelyek egyetlen levél mérésére alkalmas kamrák esetében sem könnyűek. A gondok a kamra méretével pedig hatványozottan növekednek. Az eddigi egyetlen megoldás az általunk kifejlesztett térléptékfüggő hattagú CO2 gázcseremérő kamrasorozat (Czóbel et al., 2005).
A gödöllői szünfiziológiai térléptékfüggő kamrasorozat és gyeptársulások
CO2-gázcseréjének térbeli léptékfüggése
A kamrasorozat, valamint a kamrás vegetáció és ökoszisztéma CO2-gázcsere és C-mérleg mérések technikai előkészítése és kidolgozása 1999-ben kezdődött, a rendszeres mérések pedig 2000 őszétől folynak. Az eddig ismertekhez képest szemléletében és módszertanában is új, a szünfenetikai térléptékű vizsgálatokkal párhuzamos térléptékű szünfiziológiai CO2 gázcsere mérésére alkalmas technikájú kamrasorozatot fejlesztettünk ki. Az ún. zárt rendszerű IRGA technikával működő hat darab kamra alapterülete logaritmikus skálát követ, 7,5 cm-től 480 cm-es átmérőig. A kamrákkal igen rövid időtartam (pár másodperc) alatt és széles hőmérsékleti tartományban végezhetők el a mérések. Ezzel megnyílik az út a szünfenetikai térlépték-tartományú funkcionális, beleértve a szünfiziológiai folyamatok megismeréséhez (lásd Fóti Szilvia és munkatársai tanulmányát jelen számban).
Egy másik, az ökofiziológiába ugyancsak általunk bevezetett módszer a geostatisztika alkalmazása. Ennek révén szemivariogramok segítségével – a léptékfüggő kamrasorozattal komplementer kiegészítő módon – vizsgálhatók a növényközösségek szünfiziológiai karakterisztikus léptéktartományai és a fiziológiai minimumareák (erről szintén Fóti Szilvia és munkatársai tanulmánya szól).
A növényi szünfiziológia esete is jól példázza, hogy a rendelkezésre álló módszerek jellege (más tudományoktól átvett vagy saját volta) mennyire meghatározza egy diszciplína fejlődését, amely nyilvánvalóan csak a saját módszertár kialakításával indulhat meg igazán.
Az ökofiziológiai kutatások további legújabb módszerei és irányzatai: Közülük elsőként az ún. szabadtéri kísérletes ökológiai-ökofiziológiai laboratóriumokat említeném (Tuba et al., 1998b) (1. kép). Ezekben a növénytársulások állományaival (Nagy Zoltán és munkatársai tanulmánya) vagy ökoszisztémáikkal, sőt magukkal a tájakkal folynak a kísérletek, vizsgálatok (lásd Pintér Krisztina és munkatársai munkáját). Ezen ökoszisztéma- és tájlaboratóriumokban végzett kutatások igen nagy műszer- és eszközigényű és egyben komoly fenntartási költségekkel járó hosszú távú (ún. long-term) kutatások. Azért van szükség long-term kutatásokra, mert az ökológiai jelenségeket, de még csupán az interannuális hatásokat (az egyes évek eltérő időjárásainak hatásait) is csak sokéves, évtizedes vizsgálatsorozatokból ismerhetjük meg. A kísérletekben az abiotikus és biotikus környezeti tényezőket, a növényközösségek, ökoszisztémák diverzitását, fiziognómiai és texturális szerkezetét, lomb- és gyökérarchitektúráját stb., illetve előzőek interakcióját változtatjuk meg.
Az ökofiziológiai alapfolyamatok kutatása mellett napjainkban előtérbe kerültek az éghajlattal és változásaival, valamint az ökoszisztémák, tájak használati módjával kapcsolatos kísérletes vizsgálatok is (Campbell et al., 2000), melyek színterei döntően szintén a szabadtéri kísérletes laboratóriumok. Széles körben vett geográfiai skálán az éghajlat az az állapottényező, ami leginkább meghatározza az ökoszisztéma-folyamatokat és azok szerkezetét. Az emberi tevékenység pedig gyakorlatilag az összes ökoszisztématulajdonságot és állapotot szabályozó folyamatra egyre nagyobb hatással van. Az ipari forradalom óta az emberi tevékenység olyan mértékűvé vált, hogy az egyértelműen megkülönböztethető a többi – nem emberi – hatástól, és ezért különleges figyelmet érdemel. Ezzel kapcsolatos kötetünkben Pintér Krisztina és munkatársai tanulmánya.
Új szemléletű és -keletű kutatás a filogenetikai ökofiziológia (autökofiziológiai szint), amely filogenetikai kontextusban vizsgálja a fiziológiai adaptációkat az extrém élőhelyeken. Ezzel olyan fontos és bonyolult kérdések válaszolhatók meg, mint például a C4-es fotoszintézisút vagy a N2-fixáció kialakulásának időszaka és okai. Az ökofiziológia és a filogenetika összekapcsolása új lehetőségeket nyit az organizmus biológiájának és a fiziológiai tulajdonságok evolúciójának kutatásában. Ilyen kutatás a gödöllői kiszáradástűrő növény és vegetáció projekt (inselbergökofiziológia), amely a trópusi virágos poikilohidrikus inselbergvegetáció és fajai extrém kiszáradástűrésének evolúcióját és ökofiziológiai (molekuláristól az ökoszisztémáig) alapjait vizsgálja (Oliver et al., 2000). (Erről szól kötetünkben Tuba Zoltán és munkatársai írása.)
Korunk legfontosabb gyakorlati kihívása a bioindikáció ökofiziológiai technikáinak kidolgozása táji vagy annál nagyobb tér- és időléptékű ökofiziológiai folyamatok reprezentálására. A Gödöllőn kidolgozott kriptogám transzplantációs ökofiziológiai bioindikációs eljárással a pontszerűtől a tájin és országoson át a kontinens térléptékéig végezhetők környezetállapot- (környezetterhelés-) vizsgálatok. Ráadásul maguk az alkalmazott objektumok, a mohapárnák és zuzmótelepek is szupraindividuális közösségek (populációk). Az időbeli nagy térléptéket pedig jól érzékelteti például a retrospektív bioindikációs vizsgálatok akár 100-150 éves múltat is átölelő mértéke, de a recens vizsgálatok több hónapos, féléves, éves időtartama is ezt jelzi. Mohabioindikációs munkánk révén született meg többek között hazánk első nemzeti nehézfém-légszennyezési atlasza (Ötvös et al., 2003). Ez irányú tevékenységünket Csintalan Zsolt és munkatársai tanulmánya mutatja be. Fentiek azt is érzékeltetik, hogy a növényökofiziológiában az alapkutatás és a gyakorlati alkalmazás igen közel van egymáshoz.
Kitüntetett köszönettel tartozom Fekete Gábor akadémikusnak és Bartha Sándor tudományos főmunkatársnak igen értékes és részletes szakmai véleményükért és tanácsaikért. Továbbá a CarboEurope-IP (EU 6 Framework project), OTKA 032568, PL 970518 EU-MEGARICH EU 4. RND keretprogrambeli projekt, EVK2-CT-2002-00105 GREENGRASS (EU 5 Framework project), MTA-GATE (SZIE) Globális Klímaváltozás-Növényzet kutatócsoport 10009, és MTA-SZIE Növényökológiai Kutatócsoport pénzügyi támogatásáért.
Kulcsszavak: ökofiziológia, autökofiziológia, szünfiziológia, nagy térlépték, ökoszisztéma-szénmetabolizmus, térléptékfüggés, szabadtéri kísérletes ökológiai laboratórium
Irodalom
Bartha Sándor (2004): Paradigmaváltás és módszertani forradalom a vegetáció vizsgálatában. Magyar Tudomány. 110, 12–26.
Campbell, Bruce D. – Stafford Smith, D. M. – Ash, A. J. et al. (2000): A Synthesis of Recent Global Change Research on Pasture and Rangeland Production: Reduced Uncertainties and Their Management Implications. Agriculture, Ecosystems and Environment. 82, 39–55.
Czóbel Szilárd – Fóti Sz. – Balogh J. et al. (2005): Chamber Series and Space-Scale Analysis in Grassland Vegetation. A Novel Approach. Photosynthetica. 43, 2, 267–272.
Fekete Gábor (1998): A közösségi ökológia frontvonalai. Scientia, Budapest
Juhász-Nagy Pál (1986): Egy operatív ökológia hiánya, szükséglete és feladatai. Akadémiai, Budapest
Larcher, Walter (2003): Physiological Plant Ecology. 4th ed., Springer, Berlin
Lösch, Rainer – Larcher, Walter (2000): Entwicklung und Trends der Ökophysiologie im 20. Jahrhundert. Wetter und Leben. 50, 291–336.
Mooney, H. Allan – Pearcy, R. W. – Ehleringer, J. (1987): Plant Physiological Ecology Today. BioScience. 37, 1, 18–20.
Nagy Zoltán − Pintér K. − Czóbel Sz. et al. (2007): The Carbon Budget of a Semiarid Grassland in a Wet And a Dry Year in Hungary. Agriculture, Ecosystems & Environment. 121, 21–29.
Oliver, Melvin J. – Tuba Z. – Mishler, B. D. (2000): The Evolution of Vegetative of Desiccation Tolerance in Land Plants. Plant Ecology. 151, 85–100.
Ötvös Edit – Pázmándi T. – Tuba Z. (2003): First National Survey of Atmoshperic Heavy Metal Deposition in Hungary by Analysing of Mosses. Science of Total Environment. 309, 151–160.
Tuba Zoltán – Csintalan Zs. – Szente K. et al. (1998b): Carbon Gains by Desiccation Tolerant Plants at Elevated CO2. Functional Ecology. 12, 39–44.
Tuba Zoltán – Csintalan Zs. – Nagy Z. et al. (1998a): Szünfiziologia: alapozó gondolatok és exploratív vizsgálatok egy születő növényökológiai tudományterülethez. In: Fekete Gábor (szerk.): A közösségi ökológia frontvonalai. Scientia, Budapest, 171–196.
1. táblázat

1. kép • A Gödöllői Kísérletes Növényökológiai Szabadföldi Kutatóállomás (Szent István Egyetem MKK Növénytani és Ökofiziológiai Intézet Botanikuskertje) hazánk és egyben Kelet-Európa egyetlen CO2-expozíciós szabadföldi ökológiai kísérleti laboratóriumával
<-- Vissza a 2007/10 szám tartalomjegyzékére
<-- Vissza a Magyar Tudomány honlapra
[Információk] [Tartalom] [Akaprint Kft.]