Szekeres Dóra
tanszéki mérnök
Tóth Ferenc
egyetemi docens
Szent István Egyetem Növényvédelemtani Tanszék, Gödöllő
Szénási Ágnes
egyetemi adjunktus
Szent István Egyetem Növényvédelemtani Tanszék, Gödöllő
Kádár Ferenc
tudományos segédmunkatárs
MTA Növényvédelmi Kutatóintézet, Budapest
Bevezetés
A transzgénikus növények vetésterülete első termesztési kibocsátásuk (1996) óta folyamatosan növekszik a világon, 2006-ban meghaladta a 100 millió hektárt (James, 2006). A legnagyobb területet a szója, a kukorica, a gyapot és az olajrepce tették ki, amelyek rovarrezisztens és/vagy herbicidtoleráns növények (ún. első generációs, növényvédelmi célú, azaz valamely kártevő ellen rezisztens vagy gyomirtószer-toleráns GM-növények). Az Európai Unióban jelenleg egyes ún. Bt-kukoricahibridek termesztése engedélyezett. Transzgénikus növényekből származó termékekkel (főként takarmánnyal) ma már mindannyian találkozunk.
Ugrásszerűen megnő a további, jobb termesztési, feldolgozási és felhasználási célokat (lásd jelen szám) szolgáló, genetikailag módosított növények globális termesztése, és reálisan számolhatunk az Európai Unión belüli (valamely tagállam, ideértve Magyarországot is) termesztési kibocsátásával. Vagyis a GM-növények jelenléte globálisan és a közösségen belül tény, hazai termesztési célú kibocsátással, és ennek megfelelően az abból adódó feladatokkal (ideértve a kockázatok kezelésével; részletesen később) foglalkoznunk kell.
A transzgénikus növények termesztési célú kibocsátását sok szempontból lehet és szükséges is megközelíteni, értékelni, majd ezen véleményeket, megállapításokat összevetni, ütköztetni a megfelelő gazdaság- és környezetpolitikai döntés meghozatala érdekében. A fent említett szempontok lehetnek ökonómiai elemzések különböző szintekre (egy kultúrnövény táblájára, egy gazdaság egészére, de tágabb, regionális vagy ágazati és nemzetgazdasági szintre), lehetnek környezeti kockázatokat elemző szempontok, felhasználói (takarmány- és élelmiszer-biztonsági) és etikai vagy fogyasztói szempontok is. Jelentős eltérések vannak a társadalom legkülönbözőbb rétegein belül a GM-szervezetek (növények) szükségességét, kockázatát, hatásait illetően, mely eltérések adódnak jelentős információhiányból, de adódnak eltérő érdekekből, felfogásból, sőt egyfajta bizalomhiányból is.
Cikkünkben főként hazai kutatási fejlesztési tapasztalatainkkal demonstrálva világítunk rá a környezeti kockázat becslésére és kezelésére egy kiválasztott GM-növény, az ún. Bt-kukorica példáján.
Transzgénikus növények környezeti
kockázatbecslésének alapjai
Gyakran elhangzó vélemény, főként a környezeti kockázatbecslés területén kevesebb ismerettel rendelkezőktől, hogy „még nem ismerjük a GM-növények környezeti hatását”, ami nem felel meg a valóságnak (természetesen nem tagadva azt, hogy a GM-növényekkel és azok kibocsátásának környezeti kockázatával kapcsolatos ismereteink, tudásunk és tapasztalatunk nem „teljes”, hiszen azokat lépésről lépésre szerezzük meg).
Az Európai Unióban a GM-növények környezetbe történő szándékos (így például termesztési célú) kibocsátása az ún. elővigyázatosság elvén alapszik. Ezen elvnek az a lényege, hogy egy GM-növény környezetbe történő kibocsátásának engedélyezését megelőzi az adott genetikai esemény és növény környezeti (és humán egészségügyi) kockázatelemzése. Az engedélyezési eljárás másik elve, hogy a kibocsátás fokozatosan (lépésről lépésre) történhet meg, azaz annak mértéke akkor növekedhet, ha a korábbi szinten elvégzett kockázatbecslés eredménye ezt lehetővé teszi (Directive 2001/18/EC). A továbbiakban csak a termesztési célú szándékos kibocsátással kapcsolatos kockázatbecsléssel foglalkozunk. A kibocsátási engedélyt kérőnek kötelezően benyújtandó dokumentáció tartalmát az Európai Élelmiszerbiztonsági Hatóság (EFSA – European Food Safety Authority) által kiadott Guidence Document határozza meg (EFSA Journal, 2004). A környezeti kockázatbecslés ún. lépcsőzetes módon történik a következők szerint (Wilkinson et al., 2003):
• első lépcső: a veszély meghatározása (GM-növények és termékek különböző nemcélszervezet élőlényekre gyakorolt hatásának tesztelése általában laboratóriumi, üvegházi körülmények között),
• második lépcső: táplálkozási (trofikus) szinteken keresztül gyakorolt hatás (GM-növények direkt és indirekt hatásainak vizsgálata olyan élőlényekre, amelyek nem állnak trofikus kapcsolatban a növénnyel, de a táplálékláncon keresztül érintettek lehetnek, például növényevő rovarokat fogyasztó ragadozó rovarok, általában laboratóriumi, üvegházi körülmények között),
• harmadik lépcső: szabadföldi „kitettség”-vizsgálatok, amelyek a GM-növény termesztését szimulálva, adott biotikus és abiotikus viszonyok között további információkat adnak a GM-növény esetleges nemkívánatos hatásáról, annak „kezelési” lehetőségeiről. A szabadföldi vizsgálatok rendszerint a GM-növénynek megfelelő nem GM-növénnyel történő összehasonlításokban folynak.
Az első és a második lépcső a potenciális veszélyt meghatározására, míg a harmadik lépcső az első két szint eredményei és a kitettség alapján a kockázat becslésére alkalmas.
A környezeti kockázatbecsléshez kapcsolódik a kibocsátás utáni megfigyelés (post-market monitoring), hiszen a kockázatbecslés a rendelkezésre álló tudományos eredmények alapján történik. Ezen eredmények időben hosszabb és térben nagyobb mértékű (változatos, egymástól eltérő környezeti körülmények közötti) kibocsátással folyamatosan bővülnek, és szolgálják egyrészt bármely nem várt kedvezőtlen hatás regisztrálását, másrészt folyamatos visszacsatolást a kockázatbecslés módszereinek javítására.
A kibocsátási kérelem dokumentációjának tartalmaznia kell a környezeti kockázatbecsléshez szükséges, elvégzett vizsgálatok alapján rendelkezésre álló eredményeket a befogadó szervezetről (például növény), a genetikai változásról, az előállított GM-növényről, a génbevitellel járó változásokról, tulajdonságokról, például az inzert genetikai stabilitása, lehetséges toxikus és allergén hatásokról, beltartalmi összetevőkről stb., amelyekre itt nem térünk ki (részletesen lásd: EFSA Journal, 2004).
Környezeti kockázatelemzés szempontjából egy GM-növény termesztési célú kibocsátásánál a dokumentumnak tartalmaznia kell a jellemző európai termesztési régiókra vonatkozó hatástani vizsgálatok eredményeit. Azaz a befogadó környezet szempontjából fontos, az általános kockázatbecsléshez használható eredményeket.
A környezeti kockázatbecslés szempontjából, leegyszerűsítve, mi az alábbi lehetséges hatásokat emeljük ki:
Térben: GM-növény táblája
> táblaszegély mint élőhely
> környező táblák, élőhelyek
Időben: GM-növény termesztésének éve
> következő év(ek), árvakelés, tartamhatás
Funkcionálisan: GM-növény
> rokon vad- és kultúrnövényfajok,
> célszervezet(ek) (kártevők, gyomok)
> nem célszervezetek (herbivorok, predátorok, beporzók, lebontók, stb.)
Hatás tekintetében: GM-növény
> direkt hatás:
célszervezet(ek)re hatékonyság, rezisztencia kockázata?
másodlagos kártevők gyérítése?
nem célszervezetekre toxikus hatás?
GM-növény és toxin perzisztenciája, akkumulációja a talajban?
talajfauna, mikrobiális aktivitás,
lebontó szervezetek?
vad és rokon növényfajok: átporzás,
hibridképződés ?
> indirekt hatás:
megváltozott anyagcsereösszetétel
(metabolitok),
nem célszervezetek
(zsákmány és gazda révén),
megváltozott gazdálkodási gyakorlat.
A „Bt-kukorica”
A géntechnológiával módosított rovarrezisztens növények közül jelenleg az ún. Bt-kukorica és Bt-gyapot van kereskedelmi forgalomban, termesztésben. Miután termesztését tekintve Magyarországon a kukorica jelentős növény, így ennek környezeti hatásvizsgálatára térünk ki.
Közismert, hogy az ún. Bt-kukorica a talajban élő Bacillus thuringiensis baktériumfaj valamely törzsének inszekticid fehérje (Cry toxin) termelésért felelős génjének kukorica-növénybe történő beviteléből kapta nevét. Termesztésbe először (1996) az Egyesült Államokban a kukorica egyik kártevő lepkefaja, a kukoricamoly (Ostrinia nubilalis) ellen hatékony Cry1Ab toxint termelő GM-kukorica került. A kártevő lepkefaj hernyója megrágja a növény levelét, majd annak szárába hatol be, ott táplálkozik, de megrághatja a szemeket, illetve a csutkát is. A növény sejtjeiben termelődő Cry1Ab toxin az elfogyasztott táplálékkal bejutva a hernyó emésztőrendszerébe, a középbélben lévő receptorokhoz kötődik, majd a hámsejtek membránjának károsításával a hernyó pusztulását okozza. A Cry1Ab toxin nagymértékű szelektivitást mutat, eddigi ismereteink szerint kizárólag Lepidoptera lárvák ellen hat. Különböző Bacillus thuringiensis törzsek által termelt toxinok más-más rovarcsoport ellen hatékonyak. Termesztésben jelenleg egyes Lepidoptera kártevők ellen hatékony Cry9F, Cry1F és Cry1Ac toxint és Coleoptera (Diabrotica) kártevők ellen hatékony Cry3A, Cry3Bb1, Cry34Ab1 és 35Ab1 toxint termelő kukoricahibridek találhatók meg.
A Szent István Egyetem Növényvédelemtani Tanszéke 2001 óta folytat környezeti hatásvizsgálatokat géntechnológiával módosított kukoricahibridekkel szabadföldön. Először kukoricamoly-rezisztens kukoricahibrid (EU-5 K+F „Bt-BioNoTa” projekt „Bt transzgének hatása nem célszervezet rovarok: beporzók, növényevők és ragadozók biodiverzitására”), majd később gyomirtószer-tűréssel rendelkező és/vagy lepke- és bogárkártevőkkel szemben rezisztens kukoricahibridek környezeti hatásvizsgálatát céloztuk meg.
A rovarrezisztens kukoricahibridekkel szemben az egyik fenntartás, hogy a nem célszervezet rovarokra, ízeltlábúakra is van nemkívánatos hatásuk, egyes fajok, csoportok egyedeit elpusztíthatják, vagy reprodukcióját csökkenhetti, veszélyeztetve az ízeltlábú együttes sokszínűségét, fajgazdagságát. Ezért munkacsoportunk a kockázatelemzés széles spektrumán belül a kukorica növényállományának ízeltlábú (elsősorban a talajfelszínen mozgó, vagy a talajfelszín feletti növényevő és ragadozó) együttesére végzett el hatásvizsgálatot. (Az ilyen jellegű elemzések felelnek meg a kockázatbecslés harmadik lépcsőjének.)
Miután a különböző Bt-kukoricahibridek Lepidoptera és/vagy Coleoptera kártevők ellen hatékony toxint (vagy toxinokat) termelnek, így a környezeti hatásvizsgálatban az ízeltlábú együttes mintázásában más-más nem célszervezet csoport (Lepidoptera és/vagy Coleoptera) szerepel megkülönböztetett csoportként a nemkívánatos mellékhatás megállapítása érdekében.
Cry1Ab toxint termelő hibridek:
célszervezet és más Lepidoptera fajok
Célszervezet a kukoricát károsító kukoricamoly. Kérdés, hogy a kukorica állományában jelen vannak-e, és táplálkoznak-e más Lepidoptera fajok, amelyekre a toxin hatása nemkívánatos?
A célszervezet O. nubilalis lárváit, ahogyan az várható volt, kizárólag az izogénes parcellákban találtuk meg, vagyis a Cry1Ab toxin hatékony volt a célszervezet ellen. A Cry1Ab toxint termelő kukoricában jelentősen csökkent egy, a kukorica csövén károsító másik kártevő Lepidoptera faj, a gyapottok-bagolylepke (Helicoverpa armigera) egyedszáma, kártétele (1. ábra). További kártevő Lepidoptera fajt nem mutattunk ki a felvételezések során. Más, nem kukoricanövényen táplálkozó Lepidoptera fajok lárváira akkor lehet hatással a Cry1Ab toxin, ha azok szervezetébe, például a kukorica gyomnövényei-re címerhányás időszakában lehulló pollennel az bejut. Magyarországon 187 védett lepkefaj közül ezen kockázat csak a táblaszegélyen előforduló nagy csalán (Urtica dioica) levelét fogyasztó két faj (nappali pávaszem, Inachis io, és az atalanta lepke, Vanessa atalanta) esetében jelentkezik (Darvas et al., 2004). A szerzők vizsgálatai szerint a „Bt táblák címerezett szegélysorai jelentős védelmet jelentenek a pollenszóródással szemben”. Laboratóriumi és fél-szabadföldi toxicitási eredmények gyakran túlbecsülik a Bt-kukoricának a nem célszervezet lepkefajok lárváira gyakorolt kedvezőtlen hatását, ezért a fent említett vizsgálati szintek után elengedhetetlen a konkrét szabadföldi elemzések elvégzése (Gathmann et al., 2006). Véleményünk szerint konkrét szabadföldi elemzésben (adott hibrid pollentermelő képessége, nagy csalán előfordulási gyakorisága, táblaszegély növényborítása, fajösszetétel, pollenszórás és -lerakódás mértéke a táblaszegélyben, izogénes pollencsapda-sorok stb.) alapján határozható meg a kockázatkezelés szükségessége, eszközei, módja.
Más herbivorok
A herbivor szervezetek közül a levéltetvek, kabócák és földibolhák fordultak elő leggyakrabban mind az izogénes, mind a Bt-kukoricán. Összesen tizenegy levéltetűfajt találtunk 2001–2002-ben, amelyek közül a Rhopalosi-phum padi volt domináns. Nem volt szignifikáns különbség az egyedszámot tekintve az izogénes és transzgénikus növények között, mivel a levéltetvek a floémból táplálkoznak, és nem veszik fel a toxint.
2002-ben 79 kabócafaj repült a Pherocon AM sárga ragacslapokra, ebből 63 faj fordult elő a Bt-, 49 faj az izogénes parcellákban. 2003-ban összesen 48 fajt fogtak a lapok, 36 fajt a Bt-, 37 fajt az izogénes parcellákban. A fajok közül mindkét évben a Laodelphax striatellus, Zyginidia pullula és az Empoasca solani volt domináns. Nem tudtunk kimutatni szignifikáns különbséget sem az összes egyedszám, sem az egyes felvételezési időpontok tekintetében a parcellák között. A kabócák a floémból is és a citoplazmából is vehetnek fel táplálékot, így bizonyos toxinmennyiséget.
A földibolhák közül 2002-ben 22 faj fordult elő, 17-17 faj az izogénes, illetve a Bt-parcellákon. 2003-ban 19 fajt találtunk, 17-et a Bt-, 14-et az izogénes parcellákban. 2002-ben és 2003-ban egyaránt a Phyllotreta atrát és a Phyllotreta vittulát gyűjtötték legnagyobb számban a lapok. A kabócákhoz hasonlóan a földibolháknál sem tapasztaltunk szignifikáns eltérést az egyedszámban a két kezelés között, holott e rovarok felveszik a toxint.
Az izogénes és transzgénikus kukoricában a kabóca-, illetve földibolhaegyüttesek diverzitása adott éven belül nem különbözött, a diverzitási görbék teljesen együtt futnak (2. és 3. ábra). Azaz, a domináns herbivorok tekintetében nem volt kimutatható különbség a Bt- és az izogénes kukorica között.
Predátorok
A Bt-kukorica által termelt toxin zsákmányállatra gyakorolt hatásán keresztül (denzitás, minőségbeli változás), illetve a zsákmányállat által felvett toxin hathat a kukoricában előforduló nem célszervezet predátorok abundanciájára. A kukoricában felépülő predátor együttesek egyes fajai táplálékspecialisták, mint a kizárólag levéltetveket fogyasztó katicabogarak, zöld- és barnafátyolkák illetve zengőlegyek, valamint az atkákat fogyasztó katicabogarak. Más fajok táplálékgeneralistaként nem részesítenek előnyben kizárólagos táplálékforrásként egy-egy ízeltlábú taxoncsoportot, hanem válogatás nélkül elfogyasztanak minden kisebb méretű, puhább testű állatot (például: levéltetű, atka, tripsz, rovartojások, -lárvák, -bábok), ezek közé tartoznak a futóbogarak, holyvák, tolvajpoloskák, illetve a levéltetvekkel és tripszekkel táplálkozó virágpoloskák és ragadozó tripszek. A kukoricában felépülő lombszinten aktív predátor együttesek legnagyobb részét a kizárólag levéltetveket fogyasztó afidofág fajok, míg a talajfelszínen a generalista fajok teszik ki.
2001-3 között a hatásvizsgálataink során összesen 35 növényfelszínen aktív predátor rovarfajt jegyeztünk fel a Bt- és az izogénes parcellákban. A fajszám évről évre változott. Adott évben a Bt- és az izogénes parcellák egyed- és fajszáma között nem volt szignifikáns különbség kimutatható, és az első két év során is csak 1-3 fajjal volt több a Bt-kukorica együttes fajszáma az izogénesnél. Adott éven belül a Bt- és az izogénes parcellákban igen hasonló fajösszetételű predátor együttesek épültek fel mindhárom évben (4. ábra).
Hároméves szabadföldi vizsgálatunkban az afidofág fajok közül a legjelentősebbek a katicabogarak voltak. Összesen kilenc afidofág katicabogár fajt gyűjtöttünk be. A fátyolkák közül három faj volt gyakori és domináns. Egyik évben sem tudtunk szignifikáns különbséget kimutatni az afidofág predátor fajok abundanciájában a Bt- és az izogénes parcellák között, aminek nagy valószínűséggel az az oka, hogy a levéltetvek táplálkozásuk során nem veszik fel a toxint. Vizsgálatuk mégis fontos, mivel számos predátor faj (virágtripszek, katicabogarak) fogyaszthat levéltetvek hiányában kukoricapollent, amin keresztül kapcsolatba kerülhet a toxinnal.
Az atkafogyasztó katicabogarak közül egy faj, a Stethorus punctillum volt jelen nagy számban a területen. Bár az atkák szervezetében kimutatható a toxin jelenléte, mégsem okozott szignifikáns különbséget a Bt- és az izogénes parcellák között az atkászböde abundanciájában, aminek okai az atkák bélrendszerében végbemenő deaktiválási, illetve lebontó folyamatok lehetnek (Dutton et al., 2002), vagy egyszerűen a Cry1Ab toxin rovarspecifitása (Lepidoptera lárvák). Ezen fajokon kívül számos virágpoloska faj (Orius niger, Orius majusculus és Orius minutus) volt jelen a területen, ezek a fajok levéltetű hiánya esetén atkákkal is táplálkoznak, azaz fontos fakultatív fogyasztói a tripszeknek és az atkáknak is.
A többi faj főleg generalista (futóbogarak, tolvajpoloska fajok, fülbemászók, holyvák), melyek a növényen jelen lévő kabócákkal, levéltetvekkel, poloskákkal és más kistestű rovarokkal táplálkoznak. Mindkét kukoricatípusban ugyanaz a három tolvajpoloska faj: Nabis punctatus, Nabis ferus és Nabis pseudoferus volt gyakori. A Bt- és az izogénes kukoricában gyűjtött három holyva fajból egy, a Tachyporus hypnorum volt gyakori. A vizsgálat három éve alatt összesen 58, talajfelszínen aktív futóbogár fajt gyűjtöttünk be, ezek közül mindhárom évben ugyanaz a hat faj volt domináns a Bt- és az izogénes parcellákban. Egyik évben sem tudtunk szignifikáns különbséget kimutatni a generalista predátorok abundanciájában, diverzitásában a Bt- és az izogénes kukorica között (Szekeres et al., 2006), ami nagy valószínűséggel a fajok táplálkozási módjából adódhat. Bár számos olyan zsákmányállatot fogyaszthatnak, mely tartalmazhat toxint (lepkelárvák, bábok), valószínűleg ezek csak olyan kis hányadát képezik táplálékuknak, amely nem befolyásolja az abundanciájukat.
Pókok mint generalista ragadozó fajok
Mind tudományos, mind gyakorlati szempontból fontos azon generalista ragadozó fajok vizsgálata, amelyek számára a kukorica lombozatlakó ízeltlábú együttesének majdnem minden faja potenciális zsákmányt jelent. Egyszerűbben megfogalmazva olyan ragadozót keresünk, amelyik szinte válogatás nélkül minden útjába kerülő ízeltlábút elfogyaszt a kukoricanövényen. Miért lehet érdekes egy ilyen csúcsragadozó? A transzgénikus növények várható hatása állatfajonként különböző lehet. A vizsgált fajok egy részét kedvezően, más fajokat kedvezőtlenül érinthet a transzgén jelenléte, illetve a fajok egy részénél semmilyen kimutatható hatás nem várható. A generalista csúcsragadozók populációinak elemzésével mindezek eredőjét állapíthatjuk meg, ami a taxononkénti hatásvizsgálatok értékes kiegészítője lehet. A kukoricában ilyen generalista csúcsragadozók a pókok.
Milyen jellemzőket érdemes felvételezni és mérni a pókoknál, ha a transzgén közvetett hatását keressük? Elsősorban a kérdéses faj egyedszámát, valamint az utódszámot és az elhullási arányt. Másodsorban a zsákmány faji összetételét, természetesen az egyedszámokkal, esetleg a tömeggel súlyozva. Szabadföldi mintázásunkban leggyakrabban a következő pókcsaládok egyedei fordultak elő: farkaspókok (Lycosidae), karolópókok (Thomisidae), futópókok (Philodromidae), vitorlapókok (Linyphiidae), keresztespókok (Araneidae) és a törpepókok (Theridiidae). Mivel a farkaspókok, a karolópókok és a futópókok nem hálószövő fajok, és naponta átlagosan egy–két zsákmányállatot fogyasztanak el, így táplálkozásuk nehezen követhető nyomon. A vitorlapókok és a keresztespókok hálószövő fajok ugyan, de a zsákmányt elfogyasztás közben apró darabokra morzsolják, és a maradványokat kidobják a hálóból. E sajátos táplálkozási szokásuk miatt szintén nehezen tanulmányozható a zsákmányösszetételük. Ezek alapján a törpepókokra esett a választásunk, melyek táplálkozása könnyen nyomon követhető az épen hagyott kutikulájú és a hálóba egytől egyig gondosan beszőtt zsákmányállatok segítségével. Úgy is fogalmazhatnánk, hogy a törpepókok „étlapja” hetekre visszamenőleg kiolvasható a hálótartalomból a meglepően épen maradt tetemeknek köszönhetően. A hálótartalom-vizsgálat azonban más ajándékot is rejt a kutató számára: az utódok kikelése után az üres petegubó is helyben marad. A petegubót felboncolva pontos képet kapunk mind az utódszámról, mind a kelési arányról. És mindezeket az adatokat úgy nyerjük, hogy közben a vizsgált faj populációját érintetlenül hagyjuk! Ez különösen kisparcellás kísérleteknél fontos, ahol az egyedek eltávolítása torzítaná az eredményeket. A hazai kukoricások „szuperdomináns” törpepók faja a kóró-törpepók (Theridion impressum), így hatásvizsgálatunkat is erre a fajra korlátoztuk.
Mintavételezéseink során a kóró-törpepók hálótartalom-vizsgálata széles zsákmány-spektrumot mutatott. A hálóban egyaránt megtalálhatóak voltak a levéltetvek, kabócák, levélbogarak, tripszek, növényevő és ragadozó poloskák, pattanóbogarak, földibolhák, katicabogarak, fátyolkák, zengőlegyek, hangyák, redősszárnyú darazsak, fürkészdarazsak, méhek, lepkék és hernyók. A lepkelárvák között megtalálható volt a gyapottok-bagolylepke hernyója, valamint a Bt-kukorica célszervezete, a kukoricamoly hernyója is.
A hatásvizsgálat három éve alatt sem a kifejlett kóró-törpepókok egyedszáma, sem az egyes zsákmányállat-taxonok túlnyomó többségének egyedszáma nem különbözött szignifikánsan a Bt- és az izogénes parcellákban. Ha találtunk is szignifikáns különbséget valamelyik taxon esetében valamelyik évben, a másik két év eredménye ezeket a különbségeket statisztikailag nem erősítette meg. Hasonlóképpen, az utódszámokban is mutatkozott szignifikáns különbség, amit a másik két év már nem erősített meg.
Összegezve hároméves szabadföldi eredményeinket: A vizsgált, Lepidopterák ellen hatékony Cry1Ab toxint termelő Bt-kukorica ízeltlábú együtteseire nem mutattunk ki kedvezőtlen hatást (fajszám, rajzásdinamika, egyedszám, diverzitás) a célszervezet kukoricamoly és egy másik kártevő faj, a gyapottok-bagolylepke kivételével.
Kártevő Lepidoptera és Coleoptera fajok ellen rezisztens, valamint herbicidtoleráns kukoricahibridek környezeti hatásvizsgálata
2006-ban kezdtük el korábbi tapasztalatainkra alapozva a fenti hibridek környezeti hatásvizsgálatát szabadföldön (12 kezelés × 4 ismétlés, véletlen blokk elrendezésben). Az eredmények feldolgozása, értékelése jelenleg folyik, 2007-ben megismételjük a felvételezéseket ugyanazon a helyszínen. Az előzetes adatok alapján nem valószínűsíthető kedvezőtlen hatás az ízeltlábúegyüttesre. Természetesen ez korai becslés, a feldolgozás és az idei felvételezések alapján a későbbiekben tudunk megbízható következtetéseket tenni. Munkacsoportunk véleménye szerint a GM-növények (akár transzgénikus, akár géntechnológiával módosítottak) egy új növényvédelmi lehetőséget, módszert, egy új termékelőállítási lehetőséget és egyben új kihívást is jelentenek számunkra. Fel kell arra készülnünk, hogy ha felhasználásuk gazdaságilag indokolt, környezetileg elfogadható, akkor éljünk azzal. De ehhez tapasztalatokat kell szereznünk, meg kell tanulnunk egy új kihívásra választ találni, amely tanulási folyamat a környezeti kockázatelemzésre, kockázatkezelésre is vonatkozik. Globálisan, az elmúlt tíz-tizenegy év tapasztalatairól összefoglaló elemzést találunk Olivier Sanvido, Jörg Romeis és Franz Bigler (2006) tanulmányában.
Kulcsszavak: GM-növények, Bt-kukorica, környezeti hatásvizsgálat, ízeltlábúak, biodiverzitás
Irodalom
Darvas Béla – Csóti A. – Adel, G. – Peregovits L. – Ronkay L. – Lauber É. - Polgár L. (2004): Adatok a Bt-kukoricapollen és védett lepkefajok magyarországi rizikóanaliziséhez. Növényvédelem. 40, 9, 441–449.
Dutton, Anna - Klein, H. - Romeis, J. - Bigler, F. (2002): Uptake of Bt-toxin by Herbivores Feeding on Transgenic Corn and Consequences for the Predator Chrysoperla carnea. Ecological Entomology. 27, 441–447.
EC, 2001. Directive 2001/18/EC of the European Parliment and of the Council of 12 March 2001 on the Deliberate Release into the Environment of Genetically Modified Organisms and Repealing Council Directive 90/220/EEC. Official Journal of the European Communities. L106, 1–39.
EFSA (2004): Guidance Document of the Scientific Panel on Genetically Modified Organisms for the Risk Assessment of Genetically Modified Plants and Derived Food and Feed. The EFSA Journal. 99, 1–94.
Gathmann, Achim – Wirooks, L. – Hothorn, L. – Bartsch, D. – Schuphan, I. (2006): Impact of Bt-Maize Pollen (MON 810) on Lepidopteran Larvae Living on Accompanying Weeds. Molecular Ecology. 15, 2677–2685.
James, Clive (2006): Global Status of Commercialized Biotech/GM Crops: 2006. ISAAA Brief No. 35.
Sanvido, Olivier – Stark, M.- -Romeis, J. – Bigler, F. (2006): Ecological Impacts of Genetically Modified Crops: Experiences from Ten Years of Experimental Field Research and Commercial Cultivation. Art Schriftenreihe 1. Agroscope Reckenholz-Tanikon Research Station ART, Zurich
Szekeres Dóra – Kádár F. – Kiss J. (2006): Activity Density, Diversity and Seasonal Dynamics of Ground Beetles (Coleoptera: Carabidae) in Bt- (MON810) and in Isogenic Maize Stands. Entomologica Fennica. 17, 269–275.
Wilkinson, Mike J. – Sweet, J.B. - Poppy, G. (2003): Preventing The Regulatory Log Jam; The Tiered Approach to Risk Assessments. Trends in Plant Science. 8, 208–212.
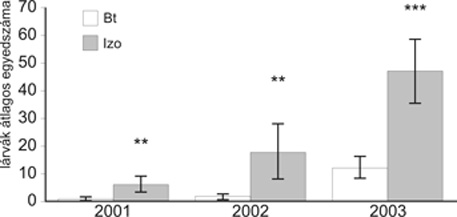
1. ábra • Helicoverpa armigera lárvák egyedszáma izogénes és transzgénikus (Bt) kukoricásokban. Szignifikáns különbség (** p<0.01 ***p<0.001) (Sóskút, 2001–2003)
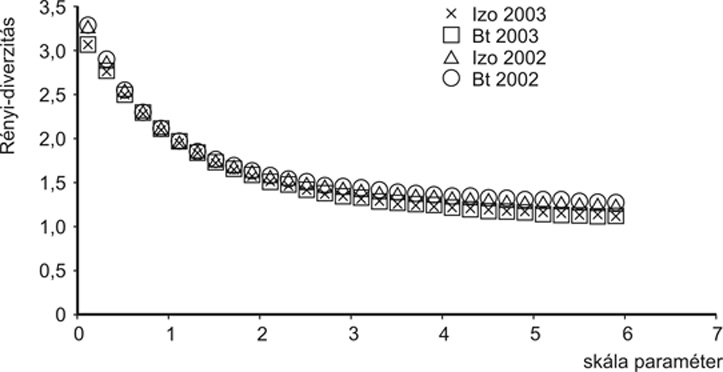 2.
ábra • Izogénes és transzgénikus
(Bt) kukoricásokban vizsgált kabócaegyüttesek
fajdiverzitási rendezése (Sóskút,
2002–2003)
2.
ábra • Izogénes és transzgénikus
(Bt) kukoricásokban vizsgált kabócaegyüttesek
fajdiverzitási rendezése (Sóskút,
2002–2003)
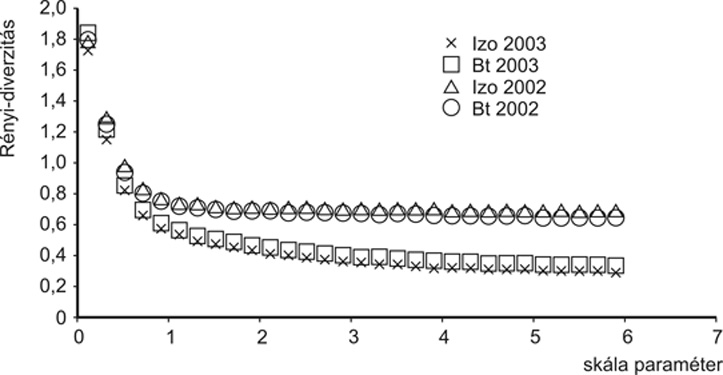 3.
ábra • Izogénes és transzgénikus
(Bt) kukoricásokban vizsgált földibolhaegyüttesek
fajdiverzitási rendezése (Sóskút,
2002–2003)
3.
ábra • Izogénes és transzgénikus
(Bt) kukoricásokban vizsgált földibolhaegyüttesek
fajdiverzitási rendezése (Sóskút,
2002–2003)
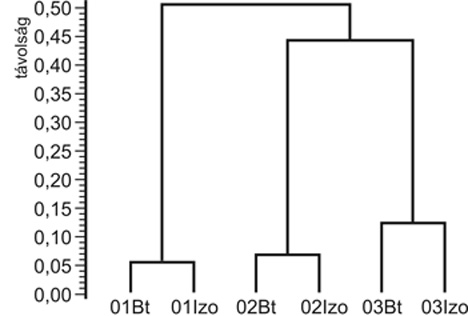 4.
ábra • Izogénes és transzgénikus
(Bt) kukoricásokban vizsgált lombszinten aktív
predátor együttesek hasonlósági
dendrogramja (Sóskút, 2001–2003)
4.
ábra • Izogénes és transzgénikus
(Bt) kukoricásokban vizsgált lombszinten aktív
predátor együttesek hasonlósági
dendrogramja (Sóskút, 2001–2003)
<-- Vissza a 2007/04 szám tartalomjegyzékére
<-- Vissza a Magyar Tudomány honlapra
[Információk] [Tartalom] [Akaprint Kft.]